


Acides nucléiques
ADN : réplication
ADN polymérase et réplisome
- Biochimie
- Chimie organique
- Bioénergétique
- Composition de la matière vivante
- Transport membranaire
- Moteurs moléculaires
- Voies de signalisation

L’élongation de la réplication de l’ADN repose sur le réplisome, un complexe multiprotéique coordonnant l’ouverture de la double hélice, la stabilisation des brins simples et la synthèse continue des nouveaux brins.
Après l’initiation de la réplication et la formation des fourches de réplication, consécutives à l’ouverture de la double hélice par l'hélicase CMG, la cellule entre dans la phase d’élongation.
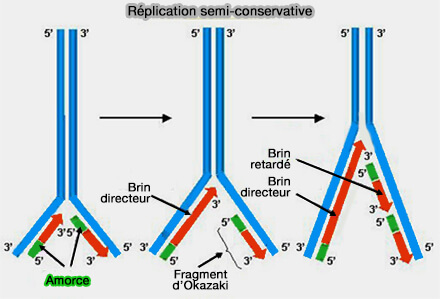
(Figure : vetopsy.fr d'après Jiménez)
{kind=link}
Cette phase d’élongation correspond à la synthèse des nouveaux brins par les ADN polymérases réplicatives organisées au sein du réplisome selon plusieurs processus successifs :
- l’amorçage de la synthèse par le complexe primase-ADN polymérase α, qui fournit l’extrémité 3'-OH indispensable au démarrage de l’élongation,
- l’élongation coordonnée du brin directeur et du brin retardé et la maturation des fragments d’Okazaki,
- les mécanismes de correction d’épreuve (proofloading) assurant la fidélité de la synthèse de l’ADN,
Ces étapes correspondent aux différents mécanismes moléculaires qui assurent la progression et l’achèvement de la réplication du génome ( terminaison de la duplication).
terminaison de la duplication).
Organisation du réplisome
Le réplisome correspond à un complexe multiprotéique dynamique assemblé au niveau de la fourche de réplication qui coordonne (Structure of a human replisome shows the organisation and interactions of a DNA replication machine 2021) :
- l’ouverture de la double hélice,
- la synthèse des nouveaux brins d’ADN,
- la progression de la fourche.
Chez les eucaryotes, ce complexe s’organise autour de l’hélicase CMG (Cdc45-MCM-GINS), qui constitue le moteur moléculaire de la fourche de réplication et assure le déroulement de l’ADN parental, et comprend plusieurs sous-ensembles fonctionnels (Caught in the act: structural dynamics of replication origin activation and fork progression 2020).

L'hélicase réplicative CMG est étudiée dans la page sur l'initiation de la réplication.
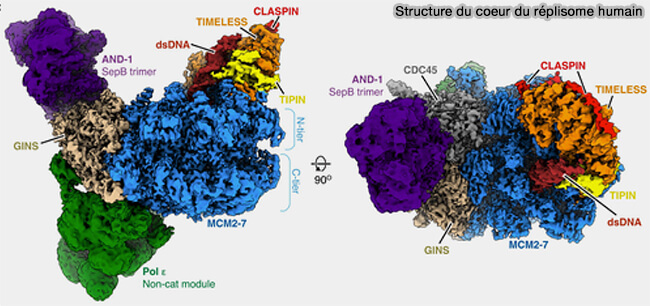
(Figure : vetopsy.fr d'après Jones et coll)
1. Les facteurs intervenant dans l’ouverture de l’ADN et la stabilisation de la fourche de réplication sont les suivants.
- Les topoisomérases relâchent les contraintes torsionnelles générées par l’ouverture de la double hélice et la progression des fourches de réplication.
- La protéine RPA (Replication Protein A) se fixe sur l’ADN simple brin généré par l’ouverture de la double hélice et en assure la stabilisation.
- Les protéines AND1/Ctf4, Mrc1/Claspin et DONSON sont associées à la fourche de réplication et contribuent à l’organisation et à la stabilité du réplisome ainsi qu’au couplage fonctionnel entre l’hélicase et les complexes de synthèse.
2. Les facteurs assurant l’initiation et l’élongation de la synthèse de l’ADN comprennent notamment.
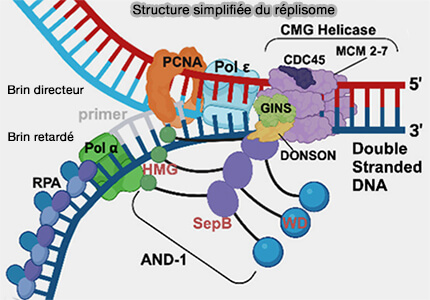
(Figure : vetopsy.fr d'après Nasheuer et coll)
a. Le complexe primase-ADN polymérase α assure l’amorçage de la synthèse en produisant les amorces ARN-ADN nécessaires au démarrage de l’élongation des nouveaux brins.
b. Les ADN polymérases réplicatives proprement dites qui comprennent :
- la polymérase ε, responsable de la synthèse du brin directeur,
- la polymérase δ, qui assure l’élongation du brin retardé.
c. Le clamp de processivité PCNA (Proliferating Cell Nuclear Antigen) forme un anneau protéique entourant l’ADN qui maintient les polymérases associées à la matrice pendant l’élongation.
Le complexe RFC (Replication Factor C) agit comme un chargeur de clamp qui ouvre transitoirement l’anneau PCNA et l’installe autour de l’ADN.
3. Les enzymes impliquées dans la maturation du brin retardé, notamment :
- RNase-H, DNA2 et Fen1/EXO1 qui assurent l’élimination des amorces et le traitement des structures en flap générées lors du déplacement de brin par la polymérase δ,
- l'ADN ligase I qui assure ensuite la ligature des fragments d’Okazaki en catalysant la formation de la liaison phosphodiester entre les fragments adjacents.
La synthèse des nouveaux brins d’ADN repose principalement sur l’activité des ADN polymérases réplicatives intégrées au réplisome.
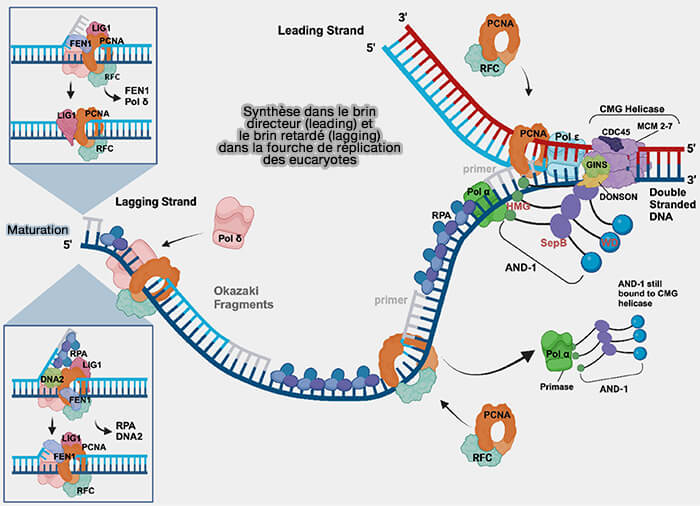
(Figure : vetopsy.fr d'après Nasheuer et coll)
ADN polymérases réplicatives
Chez les eucaryotes, plusieurs ADN polymérases, i.e.Pol α, Pol ε et Pol δ, participent à la réplication du génome nucléaire, chacune ayant une fonction spécifique dans l’organisation de la fourche de réplication (synthèse des brins directeurs et retardés).
La réplication de l'ADN mitochondrial est quant à elle assurée par une ADN polymérase spécifique, l'ADN polymérase γ (POL γ).
Remarque : d'autres ADN polymérases, notamment β, η, κ, ι, λ, μ, θ, ζ, Rev1 et PrimPol, sont principalement impliquées dans la réparation de l'ADN, la tolérance aux lésions ou d'autres fonctions spécialisées ( autres ADN polymérases).
La synthèse des nouveaux brins d’ADN est catalysée par des ADN polymérases, enzymes capables d’ajouter des désoxyribonucléotides à l’extrémité 3'-OH d’un brin d’ADN en croissance en utilisant un brin matrice complémentaire ( synthèse de l'ADN).
- La contrainte de synthèse 5' → 3' impose une organisation asymétrique lorsque les deux brins de la double hélice sont copiés simultanément.
- Dans ce complexe, les polymérases responsables de la synthèse du brin directeur et du brin retardé sont physiquement associées, ce qui permet la synthèse simultanée des deux brins parentaux.
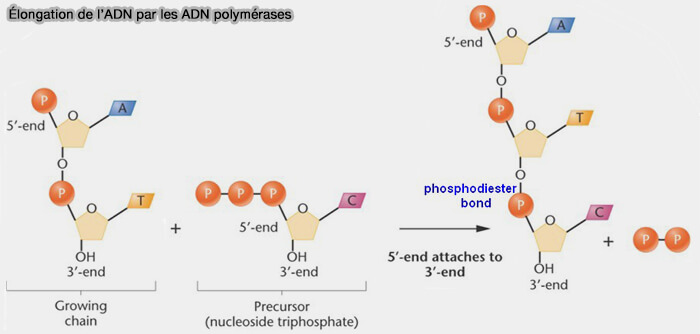
(Figure : vetopsy.fr d'après biology.stackexchange.com)
Architecture du domaine catalytique des polymérases de la famille B
Les polymérases réplicatives eucaryotes appartiennent à la famille B des ADN polymérases, caractérisée par une organisation structurale conservée et par une très grande fidélité de synthèse.
Ces enzymes possèdent une architecture tridimensionnelle typique en " main droite ", composée de trois domaines principaux qui entourent l’ADN et forment une gorge catalytique dans laquelle s’insèrent l’ADN matrice et le brin néosynthétisé.
Les autres familles d’ADN polymérases ont généralement des fonctions différentes (autres ADN polymérases).
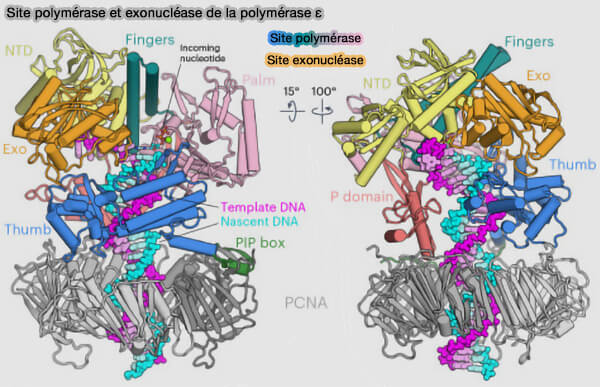
(Figure : vetopsy.fr d'après Roske et Yeeles)
1. La paume (palm) est le cœur catalytique de l’enzyme qui contient les résidus acides conservés (généralement l'aspartate ou le glutamate) qui coordonnent deux ions Mg++ indispensables à la catalyse.
a. Ces ions métalliques organisent la géométrie du site actif et stabilisent les charges négatives qui apparaissent pendant la réaction.
- Le mécanisme repose sur l’attaque nucléophile du groupement hydroxyle 3' du dernier nucléotide du brin en cours de synthèse sur le phosphate α du désoxyribonucléotide triphosphate (dNTP) entrant (incorporation des nucléotides et complémentarité des bases).
- Cette réaction forme une liaison phosphodiester entre les deux nucléotides et libère un pyrophosphate (PPi) dont l’hydrolyse rend la réaction globalement irréversible.
b. La polymérisation se fait donc exclusivement dans le sens 5' ➞ 3', tandis que le brin matrice est lu dans le sens opposé 3' ➞ 5' .
En raison de ce mécanisme, les ADN polymérases sont incapables d’initier la synthèse d’un nouveau brin de novo (direction de synthèse de l’ADN et organisation des brins).
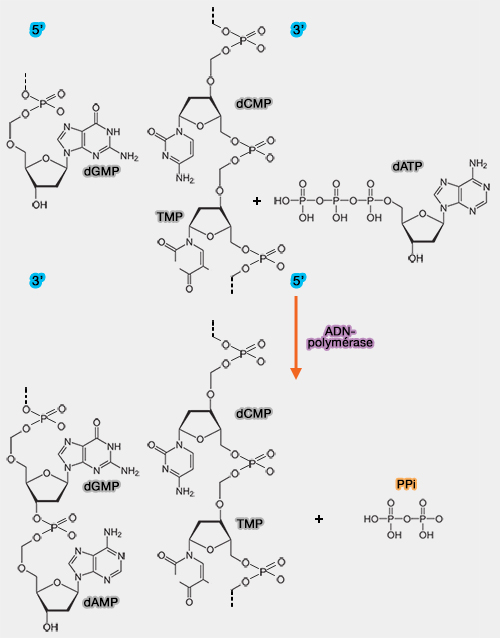
(Figure : vetopsy.fr)
2. Les doigts (fingers) constituent un domaine mobile situé au-dessus du site actif qui joue un rôle essentiel dans la sélection des désoxynucléosides triphosphates (dNTP) entrants et le contrôle géométrique contribuant à la fidélité de l’incorporation.
Lorsque le dNTP complémentaire à la base matrice diffuse dans le site polymérase, le domaine fingers subit un mouvement de fermeture qui rapproche le nucléotide du centre catalytique.
Cette fermeture aligne précisément le phosphate α du dNTP avec l’extrémité 3'-OH du brin néosynthétisé, condition nécessaire à la réaction de polymérisation.
Remarque : chez certaines ADN polymérases réplicatives, notamment Pol ε et Pol δ, l’extrémité 3' du brin néosynthétisé peut être transférée vers un site exonucléase 3' → 5' distinct du site polymérase, où le nucléotide incorrect est excisé (correction d’épreuve (proofreading) lors de la réplication de l’ADN).
3. Le pouce (thumb) forme un domaine qui entoure partiellement l'ADN double brin (dsDNA) en aval du site catalytique.
- Il établit plusieurs contacts avec le squelette phosphodiester du brin néosynthétisé et du brin matrice, ce qui stabilise la position de l’ADN dans la gorge catalytique.
- Cette interaction maintient la polymérase associée au substrat pendant l’élongation et contribue à la processivité, c’est-à-dire à la capacité de l’enzyme d’ajouter de nombreux nucléotides sans se dissocier de l’ADN.
Spécificités des polymérases réplicatives
1. La polymérase ε est la polymérase principale du brin directeur.
a. Ces caractéristiques structurales comprennent :
- une grande sous-unité catalytique POLE, qui contient le domaine polymérase, le domaine exonucléase 3'→5' et une insertion spécifique appelée domaine P (processivity domain),
- les sous-unités accessoires POLE2, POLE3 et POLE4, qui stabilisent la polymérase ε et facilitent son intégration dans l’organisation du réplisome eucaryote ;
- une interaction stable avec le complexe réplicatif CMG, qui positionne la polymérase ε immédiatement derrière l’hélicase lors de la synthèse du brin directeur.
b. Le domaine P (processivity domain) est une insertion structurale caractéristique de l’ADN polymérase ε, absente chez les polymérases réplicatives δ et α.
- Situé à proximité des sous-domaines pouce et fingers du domaine polymérase, ce segment supplémentaire forme une extension protéique qui s’approche de l'ADN double brin en aval du site catalytique.
- Ce domaine établit des contacts avec le squelette phosphodiester de l’ADN, ce qui stabilise le complexe polymérase-ADN et limite la dissociation de l’enzyme pendant l’élongation.
Cette interaction supplémentaire contribue à la forte processivité intrinsèque de Pol ε, adaptée à la synthèse continue du brin directeur dans le réplisome eucaryote.
2. La polymérase δ est la polymérase principale du brin retardé.
Ces caractéristiques structurales comprennent :
- une grande sous-unité catalytique POLD1, qui contient le domaine polymérase et le domaine exonucléase 3' → 5' assurant la correction d’épreuve,
- les sous-unités accessoires POLD2, POLD3 et POLD4, qui stabilisent le complexe polymérase δ et facilitent son interaction avec les autres composants du réplisome,
- une interaction fonctionnelle étroite avec PCNA, le clamp de processivité, qui maintient la polymérase associée à l’ADN pendant l’élongation,
- une coopération avec les enzymes de maturation des fragments d’Okazaki, notamment FEN1 et l’ADN ligase I, qui assurent respectivement l’élimination des amorces et la ligature des fragments d’ADN nouvellement synthétisés.
3. Le complexe primase-ADN polymérase α assure l’initiation de la synthèse des nouveaux brins.
Le complexe primase-ADN polymérase α est étudié dans un chapitre spécifique.
a. Ce complexe tétramérique comprend :
- Pri1 (p48), sous-unité catalytique de la primase,
- Pri2 (p58), sous-unité régulatrice de la primase,
- PolA1 (p180), sous-unité catalytique de la polymérase α,
- PolA2 (p70), sous-unité régulatrice de la polymérase.
b. Contrairement aux polymérases réplicatives ε et δ, ce complexe ne possède pas d’activité exonucléase et présente une fidélité relativement faible.
Il n’assure donc pas l’élongation principale des brins mais sert uniquement à initier la synthèse des nouveaux fragments d’ADN ( amorçage de la synthèse des nouveaux brins).
Processivité de la synthèse de l’ADN et rôle du clamp PCNA
Pour fonctionner de manière adéquate, les ADN polymérases doivent rester accrochées longtemps à l’ADN (Regulation of PCNA cycling on replicating DNA by RFC and RFC-like complexes 2019).
1. Chez les eucaryotes, cette fonction est assurée par PCNA (Proliferating Cell Nuclear Antigen), un anneau protéique trimérique qui entoure l’ADN et agit comme un clamp de processivité, maintenant les polymérases réplicatives ε et δ, associées à la matrice pendant l’élongation de la synthèse de l’ADN.
- Lors de la synthèse du brin retardé, le complexe primase-ADN polymérase α initie la synthèse d’un court segment ARN-ADN servant d’amorce, puis se dissocie rapidement. Le chargement du clamp PCNA par le complexe RFC permet alors le recrutement de la polymérase δ, qui prend le relais pour l’élongation du fragment d’Okazaki.
- PCNA forme également une plateforme d’interaction avec des facteurs de maturation des fragments d’Okazaki, comme FEN1 et l’ADN ligase I.
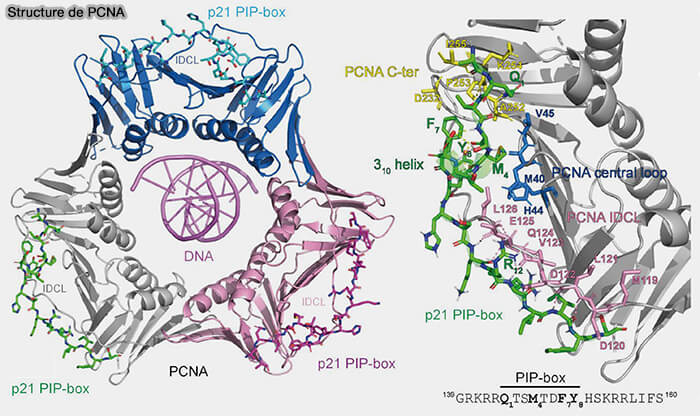
(Figure : vetopsy.fr d'après Dea Slade)
a. Chaque sous-unité possède une poche hydrophobe capable de lier un motif PIP (PCNA-interacting protein), d’environ huit acides aminés dont la séquence consensus est de type QxxΦxxΩΩ (Φ hydrophobe, Ω aromatique) ce qui permet le recrutement de plusieurs enzymes (Maneuvers on PCNA Rings during DNA Replication and Repair 2018).
- Ce motif adopte une courte hélice qui s’insère dans une poche hydrophobe située à la surface de PCNA, à proximité de la boucle IDCL, formant des interactions hydrophobes et des liaisons hydrogène stabilisant l’association protéine-PCNA.
- Cette disposition correspond au modèle “ toolbelt ”, dans lequel plusieurs enzymes sont simultanément associées au même PCNA, mais chacune occupe une sous-unité différente du trimère ( maturation des fragments d'Okasaki).
b. Le mécanisme exact complexe est très bien expliqué dans Multistep loading of a DNA sliding clamp onto DNA by replication factor (2022).
2. Le chargement de PCNA sur l’ADN est assuré par le complexe RFC (Replication Factor C), qui ouvre transitoirement l’anneau pour l’installer autour de l’ADN.
- Sur le brin retardé, le chargement de PCNA par RFC est répété à chaque amorce et permet le recrutement de la polymérase δ, responsable de l’élongation des fragments d’Okazaki.
- Sur le brin directeur, la polymérase ε est étroitement associée à l'hélicase CMG, ce qui stabilise sa progression pendant la synthèse continue du nouveau brin.
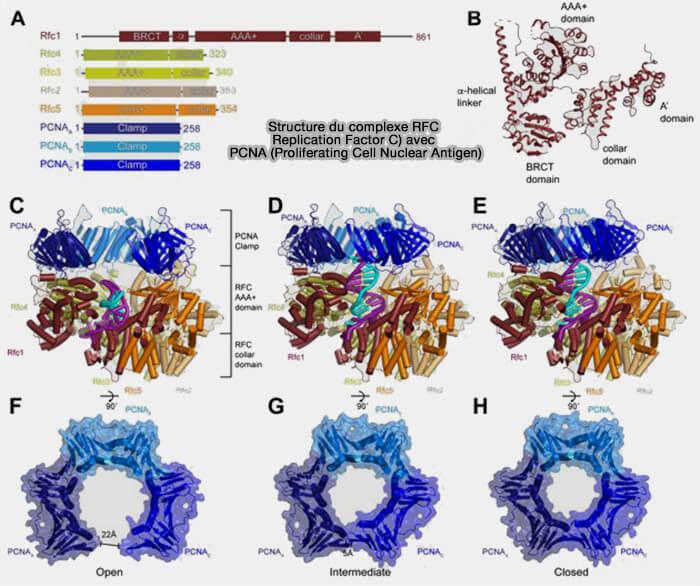
(Figure : vetopsy.fr d'après Schrecker et coll)
Contraintes topologiques et rôle des topoisomérases
Génération des contraintes torsionnelles pendant la réplication
La réplication de l'ADN génère des contraintes topologiques provoquées par l’ouverture de la double hélice et la progression des fourches de réplication et les topoisomérases jouent un rôle essentiel dans leurs résolutions.
1. Lorsque l’hélicase CMG ouvre la double hélice au niveau de la fourche de réplication, elle sépare progressivement les deux brins parentaux en rompant les liaisons hydrogène entre les bases, ce qui modifie la topologie de l’ADN (Closing the DNA replication cycle: from simple circular molecules to supercoiled and knotted DNA catenanes 2019).
a. Celle-ci est décrite par trois paramètres liés par la relation Lk = Tw + Wr.
- Le linking number (Lk) correspond au nombre total d’enlacements entre les deux brins et reste constant tant que l’ADN n’est pas coupé.
- Le twist (Tw) représente le nombre de tours de la double hélice.
- Le writhe (Wr) correspond aux superenroulements de la molécule dans l’espace.
Remarque : les schémas de topologie de l’ADN utilisent souvent des molécules circulaires, car le linking number (Lk) n’est défini et conservé que pour un ADN topologiquement fermé.
- Dans un ADN linéaire libre, les extrémités peuvent tourner et relâcher la torsion, ce qui empêche de définir correctement les relations entre Lk, twist (Tw) et writhe (Wr).
- Les chromosomes eucaryotes étant organisés en domaines topologiquement contraints, ils se comportent localement comme des segments fermés, ce qui permet d’appliquer les mêmes principes.
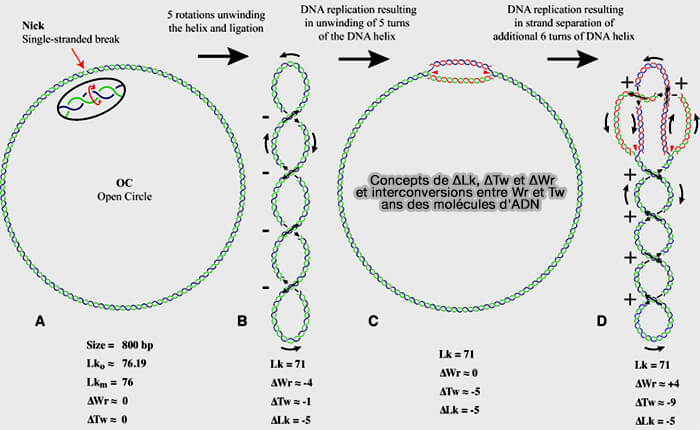
(Figure : vetopsy.fr d'après Schrecker et coll)
b. Lorsque l’hélicase ouvre l’ADN à la fourche de réplication, le nombre de tours (Tw) de la double hélice diminue localement car l’ADN est très long, associé à des histones, organisé en chromatine ancrée dans des domaines chromosomiques et la molécule ne peut donc pas tourner librement.
Comme Lk ne peut pas changer sans coupure de l’ADN, cette variation est compensée par une augmentation des superenroulements (Wr).
c. Les tours d’hélice deviennent plus serrés et on parle alors de surenroulement positif.
- Autrement dit, derrière la fourche, l’ADN est déjà déroulé et simple brin, donc il ne peut pas stocker de torsion hélicoïdale.
- Devant la fourche, l’ADN reste en double hélice et absorbe la contrainte sous forme de surenroulements positifs.
d. Si ces contraintes torsionnelles n’étaient pas dissipées, elles entraîneraient un blocage rapide de la progression de l’hélicase CMG et des ADN polymérases.
Rôles des topoisomérases dans la levée des contraintes
1. Les topoisomérases interviennent pour modifier le Lk en coupant transitoirement l’ADN, en introduisant transitoirement des coupures strictement contrôlées dans la molécule, ce qui permet de relâcher les tensions accumulées lors de la réplication tout en conservant l’intégrité de l’ADN (Immunodetection of human topoisomerase I-DNA covalent complexes 2016).
a. Lors de la réaction, un résidu tyrosine du site actif de l’enzyme attaque la liaison phosphodiester de l’ADN et forme un intermédiaire covalent enzyme-ADN.
- L’enzyme reste donc temporairement attachée à l’extrémité de l’ADN coupé, ce qui empêche toute dissociation incontrôlée des brins.
- Cette coupure permet alors à la molécule d’ADN de se détordre et de relâcher les surenroulements accumulés.
b. Une fois la torsion relâchée, la réaction se produit en sens inverse :
- L’extrémité 3'-OH ou 5'-OH libre de l’ADN attaque la liaison phosphotyrosine enzyme-ADN et reforme la liaison phosphodiester normale, ce qui religature la molécule.
- L’enzyme se détache alors et l’ADN retrouve sa continuité structurale.
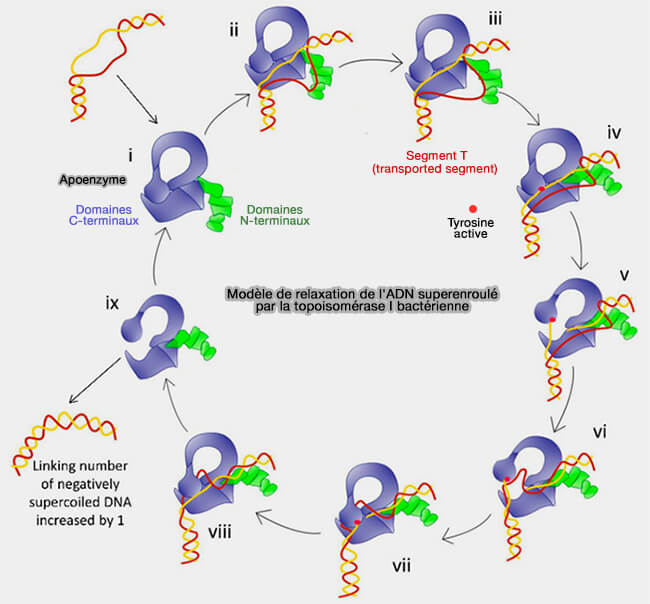
(Figure : vetopsy.fr d'après Dasgupta et coll)
2. On distingue deux grandes classes de topoisomérases.
a. Les topoisomérases de type I coupent transitoirement un seul brin d’ADN (Mechanism of Type IA Topoisomerases 2020).
- Cette coupure permet la rotation du brin intact autour du brin coupé et ce mécanisme ne nécessite généralement pas d’hydrolyse d’ATP.
- Les topoisomérases I peuvent former une liaison 3'-phosphotyrosine (Topo IB eucaryote) ou une liaison 5'-phosphotyrosine (Topo IA).
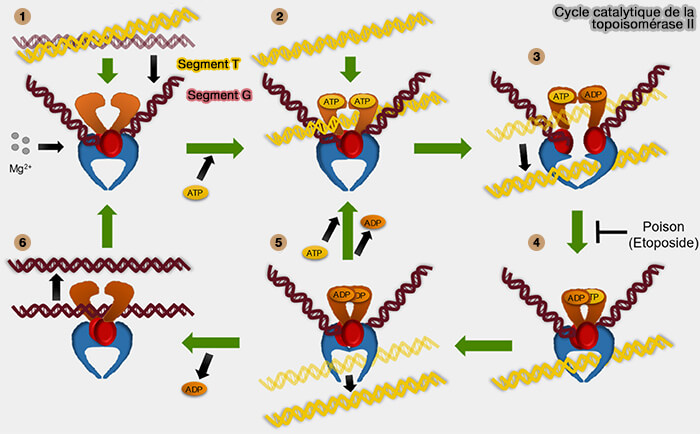
(Figure : vetopsy.fr d'après Gibson et Deweese)
Remarque : certaines topoisomérases de type IA eucaryotes, en particulier TOPIIIα, n’agissent pas principalement dans la relaxation des supertours de l’ADN mais participent à des processus de recombinaison (dissolution topologique dans la recombinaison homologue).
b. Les topoisomérases de type II coupent transitoirement les deux brins de la double hélice et l’enzyme agit comme une pince moléculaire contrôlant l’ouverture de l’ADN double brin (Structural and Biochemical Basis of Etoposide-Resistant Mutations in Topoisomerase IIα 2022).
a. L’enzyme se fixe d’abord sur un segment d’ADN double brin appelé segment G (gate segment), puis coupe simultanément les deux brins de cette molécule.
- Cependant, chaque extrémité de l’ADN reste covalemment liée à un résidu tyrosine de l’enzyme, ce qui maintient les extrémités sous contrôle et empêche toute rupture irréversible de la molécule.
- L’enzyme peut alors écarter légèrement les deux extrémités qu’elle maintient, créant une ouverture transitoire dans la double hélice.
b. À travers cette ouverture, un autre segment d’ADN double brin appelé segment T (transported segment) peut être déplacé grâce à l’énergie fournie par l’hydrolyse de l’ATP.
c. Une fois ce passage effectué, l’enzyme rapproche les extrémités maintenues et reforme la liaison phosphodiester, ce qui restaure la continuité de l’ADN.
Le rôle de la topoisomérase IIα, enzyme essentielle à la séparation des chromatides soeurs, est étudié dans un chapitre spécifique.
Remarque : outre les topoisomérases de type I et II impliquées dans la gestion de la topologie de l’ADN lors de la réplication et de la transcription, d’autres enzymes apparentées existent dans certains organismes ( reconnaissance des cassures double brin).
Amorçage de la synthèse des nouveaux brins
La synthèse des nouveaux brins d’ADN ne peut pas commencer directement par l’action des polymérases réplicatives, car ces enzymes ne sont capables d’allonger qu’une extrémité 3'-OH préexistante.
L’initiation de la synthèse nécessite donc la formation d’une amorce d’ARN synthétisée par le complexe primase-ADN polymérase α, qui fournit le point de départ indispensable à l’élongation du brin directeur et du brin retardé.
BiochimieChimie organiqueBioénergétiqueAcides nucléiquesNucléotidesADNStructureSynthèseOrganisation du génomeRéplicationTranscriptionÉpigénétiqueRéparation de l'ADNARNChromatineNucléosomesHistonesChromosomesProtidesGlucidesLipidesEnzymesCoenzymesVitaminesHormonesComposés inorganiquesTransport membranaireMoteurs moléculairesVoies de signalisation