


Acides nucléiques
ADN : réparation
des cassures double brin
Recombinaison homologue (HR)
Voies de réparation après formation de la D-loop
- Biochimie
- Chimie organique
- Bioénergétique
- Composition de la matière vivante
- Transport membranaire
- Moteurs moléculaires
- Voies de signalisation

Les systèmes de réparation de l’ADN reposent sur des ensembles enzymatiques spécialisés (nucléases, polymérases et ligases) qui détectent les lésions, excisent les segments endommagés et restaurent l’intégrité du génome.
La D-loop, formée après l’invasion de brin et la synthèse initiale d’ADN à partir de la chromatide sœur, constitue l’intermédiaire central de la recombinaison homologue et peut ensuite évoluer selon deux voies principales de réparation.
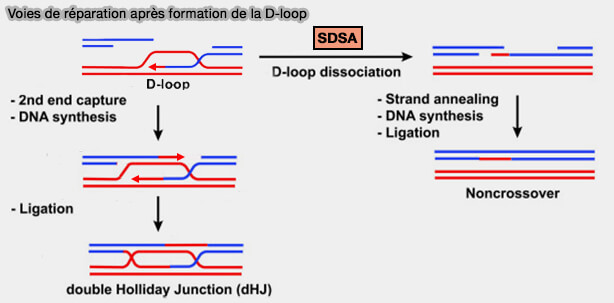
Voies de réparation après formation de la D-loop
Après la formation de la D-loop et synthèse d’ADN, deux voies principales peuvent être suivies par l’intermédiaire de recombinaison.
1. Dans la voie SDSA (Synthesis-Dependent Strand Annealing), dominante dans les cellules somatiques, la D-loop est démantelée après la synthèse initiale.
a. Après l’invasion du brin par le filament RAD51 et la formation de la D-loop, l’extrémité 3′ envahissante est allongée par les polymérases réplicatives, principalement la polymérase δ p associée au clamp PCNA et au chargeur RFC.
b. Lorsque la synthèse a recopié une portion suffisante de la séquence homologue, la D-loop est dissociée par des hélicases de recombinaison, notamment BLM, qui déstabilisent l’intermédiaire d’invasion de brin et libèrent le brin nouvellement synthétisé.
- Le brin néosynthétisé se détache alors de la matrice homologue et s’apparie avec l’autre extrémité résectée de la cassure grâce aux régions complémentaires générées par la synthèse d’ADN.
- Les éventuels segments simple brin excédentaires sont éliminés par des nucléases de structure, notamment FEN1, les lacunes sont ensuite comblées par les ADN polymérases, principalement la polymérase δ, et la réparation est achevée par la ligature des brins, généralement assurée par l'ADN ligase I.
c. Dans la voie SDSA, la seconde extrémité de la cassure n’envahit pas la chromatide sœur et se réapparie directement avec le brin nouvellement synthétisé après la dissociation de la D-loop, ce qui empêche la formation de doubles jonctions de Holliday et conduit exclusivement à des produits non-crossover.
(Figure : vetopsy.fr d'après Daley et coll)
2. Dans la voie de capture du second brin (second-end capture), la seconde extrémité de la cassure possède aussi un brin simple 3' qui peut s’apparier avec le brin déplacé de la D-loop, étape appelée capture du second brin (second-end capture).
a. Après extension de la synthèse d’ADN et ligature des extrémités, il se forme un intermédiaire de recombinaison contenant deux jonctions de Holliday (double Holliday junction) caractérisé par :
- deux points de croisement des brins,
- deux régions d’ADN hétéroduplex.
b. Ces jonctions peuvent ensuite être traitées soit par dissolution topologique, soit par résolution nucléolytique, ce qui détermine la formation d’un produit crossover ou non-crossover.
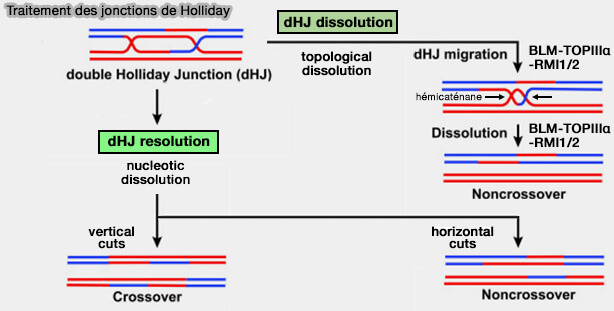
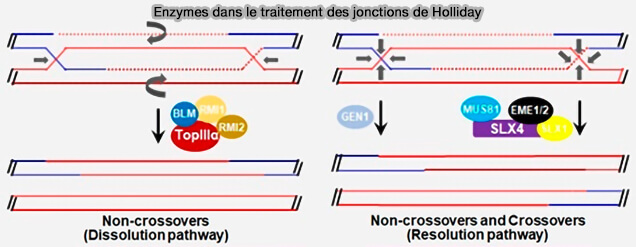
Traitement des jonctions de Holliday
Après la formation d’une double jonction de Holliday (double Holliday junction, DHJ), l’intermédiaire de recombinaison doit être éliminé afin de séparer les deux molécules d’ADN.
- la dissolution topologique (dHJ dissolution)
- la résolution nucléolytique (dHJ resolution)
(Figure : vetopsy.fr d'après Daley et coll)

Vous pouvez lire : DNA Holliday Junction: History, Regulation and Bioactivity (2022).
Dissolution topologique (dHJ dissolution)
Dans la dissolution topologique, voie majoritaire en mitose, les deux jonctions de Holliday migrent l’une vers l’autre jusqu’à converger (convergent branch migration).
La dissolution topologique des dHJ est réalisée par le complexe BTR (BLM-TOPIIIα-RMI1-RMI2), appelé aussi dissolvasome.
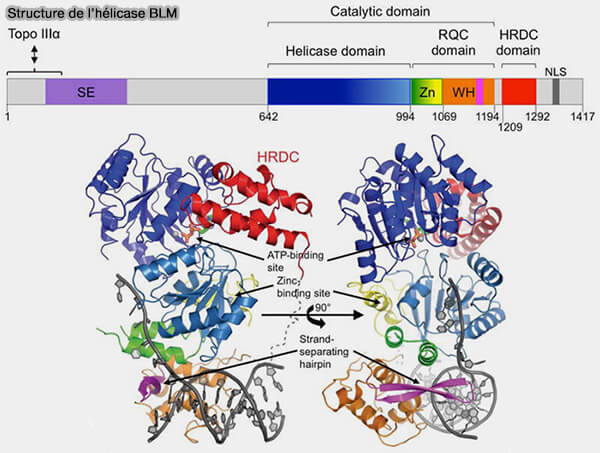
1. Ce déplacement des jonctions est catalysé par l’hélicase BLM (Bloom syndrome protein), qui agit comme une hélicase de migration de branche et déplace progressivement les points de croisement le long des molécules d’ADN sans rompre les brins (Bloom’s Syndrome: Why Not Premature Aging? A comparison of the BLM and WRN helicases 2018).
a. BLM appartient à la famille RecQ des hélicases ADN et possède une organisation modulaire adaptée à la reconnaissance des intermédiaires de recombinaison.
- Son domaine hélicase ATP-dépendant assure l’activité motrice nécessaire au déplacement des jonctions de Holliday.
- Le domaine RQC participe à la liaison à différentes structures d'ADN, notamment les jonctions de recombinaison et certaines structures secondaires
- Le domaine HRDC contribue à la spécificité de reconnaissance de substrats spécifiques et à l'interaction avec certaines structures d'ADN impliquées dans la recombinaison homologue et la réparation.
(Figure : vetopsy.fr d'après de Renty et Ellis)
b. L’hélicase BLM se fixe sur la jonction et utilise l’ATP pour déplacer le point de croisement et le déplacement correspond à un échange progressif des paires de bases, ce qui fait glisser la jonction le long de l’ADN.
c. La migration convergente rapproche les deux jonctions de Holliday jusqu’à leur conversion en un intermédiaire topologique appelé hémicaténane, dans lequel les deux molécules d’ADN restent entrelacées.
2. Cet intermédiaire est ensuite résolu par la topoisomérase TOPIIIα, qui réalise une réaction de décaténation en coupant transitoirement un brin d’ADN, en faisant passer l’autre brin à travers la coupure puis en religuant l’ADN (Structural and mechanistic insight into Holliday junction dissolution by Topoisomerase IIIα and RMI1 2015).
Ce mécanisme permet de séparer les deux molécules d’ADN sans clivage des jonctions de Holliday.
(Figure : vetopsy.fr d'après Wang et coll)
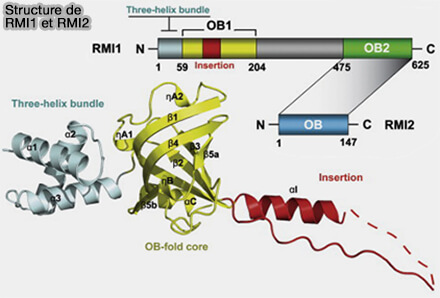
3. L’ensemble de ces réactions est assuré par le complexe dissolvasome constitué de l’hélicase BLM, de la topoisomérase TOPIIIα et des protéines accessoires RMI1 et RMI2, qui stabilisent le complexe et stimulent l’activité de la topoisomérase (Crystal Structures of RMI1 and RMI2, Two OB-Fold Regulatory Subunits of the BLM Complex 2010).

La dissolution topologique conduit exclusivement à des produits non-crossover et constitue la voie majoritaire de traitement des doubles jonctions de Holliday dans les cellules somatiques.
Dissolution nucléolytique (dHJ résolution)
Dans la résolution nucléolytique, les jonctions de Holliday sont clivées par des endonucléases spécialisées appelées résolvases (Holliday Junction Resolvases 2014).
1. GEN1, la résolvase canonique des jonctions de Holliday intactes, introduit deux coupures coordonnées sur des brins opposés de la jonction.
- Ces incisions sont réalisées à des positions équivalentes sur les deux duplex, ce qui génère deux extrémités ligaturables et conduit directement à la séparation des deux molécules d’ADN.
- GEN1 agit principalement en fin de mitose ou en télophase après l’entrée de GEN1 dans le noyau lors de la rupture de l’enveloppe nucléaire lorsque les intermédiaires de recombinaison persistent.
2. Dans la résolution des jonctions partiellement migrées ou déjà entaillées (nick), les nucléases associées à SLX4 interviennent selon un mécanisme différent (Coordinated Actions of SLX1-SLX4 and MUS81-EME1 for Holliday Junction Resolution in Human Cells 2013).
a. Le complexe nucléasique SLX1-SLX4, dans lequel SLX4 agit comme une plateforme recrutant et positionnant la nucléase SLX1, peut introduire une première coupure dans la jonction de Holliday, créant une structure asymétrique.
Cette première incision transforme la jonction de Holliday intacte en une jonction entaillée (nick), qui devient alors un substrat privilégié pour la nucléase MUS81-EME1, endonucléase structure-spécifique qui agit préférentiellement sur des structures d’ADN déjà entaillées ou ramifiées.
(Figure : vetopsy.fr d'après Song et coll)
b. MUS81-EME1 réalise ensuite une seconde coupure sur le brin opposé, ce qui permet la séparation finale des molécules d’ADN.
c. Ce mécanisme séquentiel explique pourquoi le complexe SLX-MUS agit comme une résolvase composite, la plateforme SLX4 coordonnant l’action successive de SLX1 et de MUS81-EME1.
- Ce complexe se forme surtout en fin de mitose lorsque des intermédiaires de recombinaison persistent ou lorsque la dissolution par le dissolvasome BLM-TOPIIIα-RMI1-RMI2 n’a pas abouti et c’est donc une voie de secours pour éliminer les structures de recombinaison avant la séparation des chromatides.
- Ce mécanisme est régulé par les kinases mitotiques (CDK1, Plk1) qui activent MUS81 à la transition G2/M.
(Figure : vetopsy.fr d'après Feng et coll)
Remarque : la protéine d’échafaudage SLX4 intervient également dans la voie de réparation SSA (single-strand annealing, en recrutant la nucléase XPF-ERCC1.
Dans certains travaux, l’ensemble des nucléases associées à la plateforme SLX4, comprenant SLX1-SLX4, MUS81-EME1 et XPF-ERCC1, est désigné sous le terme de complexe SMX.
Recombinaison homologue en mitose et méiose
1. Dans les cellules somatiques, la recombinaison homologue vise principalement à restaurer l’intégrité du génome sans produire d’échanges chromosomiques et permet d’éliminer les jonctions de Holliday restantes avant la séparation des chromatides soeurs.
Les crossovers peuvent en effet entraîner une perte d’hétérozygotie ou des réarrangements chromosomiques.
- La cellule privilégie donc la dissolution des jonctions de Holliday par le dissolvasome BLM-TOPIIIα-RMI1-RMI2.
- Lorsque cette dissolution est impossible, les résolvases nucléolytiques, SLX-MUS (SLX1-SLX4-MUS81-EME1) et GEN1, interviennent selon une hiérarchie temporelle.
2. En méiose, la recombinaison homologue remplit une fonction différente.
- Les échanges entre chromosomes homologues sont nécessaires pour assurer la formation de chiasmas, qui maintiennent les homologues associés et garantissent leur bonne ségrégation lors de la première division méiotique.
- Certaines doubles jonctions de Holliday sont donc stabilisées et orientées vers la formation de crossovers pour générer des échanges chromosomiques indispensables à la diversité génétique et à la ségrégation correcte des chromosomes homologues.
Les crossovers de la méiose sont étudiés dans un chapitre spécifique.
a. Ce processus implique notamment :
- le complexe MutSγ1 formé par MSH4 et MSH5, qui stabilise les intermédiaires de recombinaison homologue,
- RNF212/HEI10 qui participent à la sélection et à la maturation des sites destinés à devenir des crossing-over,
- le complexe MutLγ (MLH1-MLH3), une résolvase méiotique spécialisée qui catalyse la résolution des jonctions de Holliday en produits crossover.
b. Les résolvases mitotiques, principalement MUS81-EME1, ne disparaissent pas pour autant en méiose et interviennent encore pour :
- les crossovers de classe II,
- les intermédiaires HR non traités par MutLγ.
3. Dans la résolution nucléolytique, l’orientation des coupures détermine la nature du produit final.
- Si les coupures sont réalisées dans le même plan (vertical cuts), les chromosomes échangent leurs bras, ce qui produit un crossing-over, phénomène particulièrement fréquent lors de la méiose.
- Si les coupures sont réalisées dans des plans opposés (horizontal cuts), la réparation conduit à un produit non-crossover sans échange chromosomique.
Remarque : les régions d’hétéroduplex formées pendant la recombinaison peuvent contenir des mésappariements.
- Lorsque les deux molécules d’ADN homologues ne sont pas parfaitement identiques, l’appariement entre les brins peut générer des bases non complémentaires dans la région d’hétéroduplex.
- Ces mésappariements sont corrigés par le système de réparation des mésappariements (MMR).
Réparation des cassures double brin par jonction des extrémités
La recombinaison homologue constitue une voie de réparation très fidèle des cassures double brin, mais elle dépend de la présence d’une matrice homologue, généralement la chromatide sœur.
D’autres mécanismes permettent également de réparer ces cassures en reliant directement les extrémités d’ADN ou en utilisant de courtes régions d’homologie, processus regroupés sous les voies de réparation par jonction des extrémités d’ADN (NHEJ, SSA, MMEJ/TMEJ).
BiochimieChimie organiqueBioénergétiqueAcides nucléiquesNucléotidesADNStructureSynthèseOrganisation du génomeRéplicationTranscriptionÉpigénétiqueRéparation de l'ADNARNChromatineNucléosomesHistonesChromosomesProtidesGlucidesLipidesEnzymesCoenzymesVitaminesHormonesComposés inorganiquesTransport membranaireMoteurs moléculairesVoies de signalisation