


Nucléotides : structure
Bases azotées
Organisation dans les acides nucléiques
- Biochimie
- Transport membranaire
- Moteurs moléculaires
- Voies de signalisation

L’organisation des bases azotées dans l’ADN et l’ARN repose sur l’appariement complémentaire, l’empilement des bases et plusieurs interactions stabilisant la structure des acides nucléiques.
(Figure : vetopsy.fr d'après OpenStax)
{kind=link}
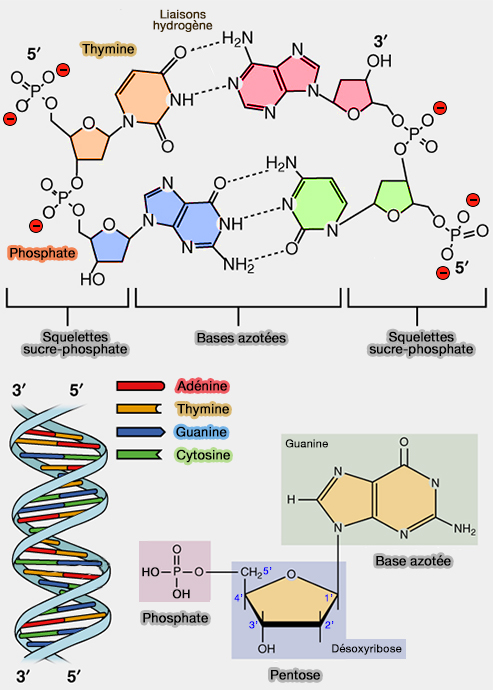
Agencement des bases dans l'ADN
L’agencement des bases azotées dans les acides nucléiques suit des règles précises, dictées à la fois par leurs dimensions, leur planarité, et leur capacité à former des liaisons hydrogène spécifiques.
Appariement complémentaire des bases
1. Dans l’ADN, les bases sont appariées selon la complémentarité des bases décrite par la règle de Chargaff :
- l’adénine (A) s’apparie avec la thymine (T) par deux liaisons hydrogène, A=T,
- la cytosine (C) s’apparie avec la guanine (G) par trois liaisons hydrogène, G≡C.
2. Cet appariement respecte une distance inter-base constante (~1,08 nm) et assure une géométrie régulière de la molécule d’ADN en double hélice droite de type B ( paires de bases dans l'ADN).
paires de bases dans l'ADN).
- La complémentarité repose sur la disposition des donneurs et accepteurs de liaisons hydrogène ainsi que sur les dimensions respectives des bases puriques et pyrimidiques ( propriétés géométriques des bases).
- La stabilité de la double hélice dépend également en partie de la proportion de paires G≡C, qui forment trois liaisons hydrogène et présentent des interactions d’empilement plus favorables que les paires A=T.
3. Les mésappariements de bases correspondent à l’incorporation d’un nucléotide non complémentaire lors de la réplication de l’ADN.
Ils peuvent générer localement des déformations structurales telles que des bulges, correspondant à l’extrusion d’un ou plusieurs nucléotides non appariés sur un seul brin, induisant une asymétrie et une distorsion de la double hélice.
a. La plupart de ces erreurs sont immédiatement corrigées par les ADN polymérases réplicatives,
- soit par la sélection correcte du nucléotide lors de l’incorporation,
- soit par leur activité de relecture exonucléase 3' → 5' (proofreading) qui élimine les bases mal appariées.
b. Les erreurs qui échappent à ces mécanismes sont ensuite corrigées par le système de réparation des mésappariements (MMR), qui détecte la distorsion locale de la double hélice et remplace le segment du brin néosynthétisé contenant l’erreur.
Polarité des brins et orientation antiparallèle (3'–5')
L’ADN est constitué de deux brins enroulés en double hélice, chacun formé d’une succession de nucléotides reliés par des liaisons phosphodiester.
1. Chaque brin possède une polarité structurale :
- une extrémité 5′, portant un groupe phosphate libre,
- une extrémité 3′, portant un groupe hydroxyle ($\ce{-OH}$) libre sur le carbone 3′ du désoxyribose.
2. Les deux brins de l’ADN sont antiparallèles, c’est-à-dire orientés en directions opposées :
- l’un dans le sens 5' ➞ 3′,
- l’autre dans le sens 3′ ➞ 5′.
3. Cette organisation est essentielle à plusieurs niveaux (antiparallélisme des brins).
Interactions stabilisatrices de la double hélice
Outre les liaisons hydrogène entre bases complémentaires, d’autres forces stabilisent la structure des acides nucléiques.
Interactions π-π : empilement vertical des bases
Les interactions π-π désignent les forces attractives entre les nuages d’électrons π délocalisés des cycles aromatiques des bases azotées.
- Les électrons π correspondent aux électrons impliqués dans les doubles liaisons des cycles aromatiques. Lorsque ces doubles liaisons sont conjuguées, les électrons π peuvent se délocaliser sur l’ensemble du cycle, ce qui confère aux bases azotées leur caractère aromatique et leur planéité.
- Le caractère aromatique des bases résulte d’un système π conjugué respectant la règle de Hückel (4n + 2 électrons π), comme dans de nombreuses molécules aromatiques biologiques.

Dans de nombreux cas, la contribution énergétique de l’empilement des bases est au moins aussi importante, voire supérieure, à celle des liaisons hydrogène pour la stabilité globale des duplex d’acides nucléiques.
1. Dans l’ADN et l’ARN, les bases sont quasi-planaires et empilées les unes au-dessus des autres le long de l’axe de l’hélice, séparées par une distance moyenne d’environ 0,34 nm entre deux bases successives ( propriétés géométriques des bases).
Cet empilement régulier permet l’établissement d’interactions π-π entre les cycles aromatiques adjacents.
- Ces interactions ne correspondent pas à des liaisons covalentes, mais à des interactions de van der Waals associées à la polarisation des nuages électroniques π.
- Elles sont de courte portée et cumulatives : la répétition de l’empilement des bases le long du brin confère une contribution majeure à la stabilité de la structure.
2. Ces interactions π-π jouent ainsi un rôle central dans la stabilisation de la double hélice de l’ADN, en complément des liaisons hydrogène reliant les bases complémentaires.
Dans les régions riches en paires G≡C, l’empilement des bases est généralement plus favorable, ce qui contribue à une stabilité thermique plus élevée et à une température de fusion plus importante.
3. Ces interactions ne sont pas des liaisons covalentes, mais des forces de van der Waals renforcées par la polarisation des cycles aromatiques.
- Elles sont directionnelles, courtes et cumulatives : plus les bases sont empilées de manière régulière, plus la stabilité de la double hélice est grande.
- Elles stabilisent fortement la structure de l’ADN, en complément des liaisons hydrogène.
- L’empilement π est également favorisé par l’exclusion de l’eau, renforçant les interactions hydrophobes entre bases.
4. Dans l’ARN, ces interactions participent également à la stabilisation des structures secondaires et tertiaires, notamment dans les hélices internes et dans certaines structures complexes telles que les boucles ou les pseudonœuds.
Contribution hydrophobe à la stabilité de la double hélice
Les bases azotées sont globalement peu polaires en comparaison avec les groupes phosphate du squelette pentose-phosphate, qui portent des charges négatives et interagissent fortement avec l’eau.
1. Dans la double hélice d’ADN, cette différence de polarité conduit à une organisation structurale caractéristique.
- Les bases azotées sont regroupées au centre de l’hélice et empilées les unes au-dessus des autres,
- le squelette pentose-phosphate reste exposé au milieu aqueux.
2. Cette disposition limite les interactions défavorables entre les surfaces aromatiques relativement hydrophobes des bases et les molécules d’eau.
- L’exclusion du solvant aqueux n’est pas active mais résulte de l’effet hydrophobe, favorisant l’empilement des bases et renforce les interactions π-π entre cycles aromatiques.
- Les bases sont protégées du solvant au cœur de la structure, alors que les groupements phosphate hydrophiles interagissent avec le milieu aqueux et les cations environnants.
Interactions ioniques
Les acides nucléiques possèdent un squelette pentose-phosphate chargé négativement, en raison des groupements phosphate ionisés (-PO4--).
Ces charges créent une forte répulsion électrostatique entre les brins d’ADN ou les segments repliés d’un ARN.
La stabilité des structures d’acides nucléiques dépend donc en partie de la présence de cations capables de compenser cette répulsion.
1. Les ions positifs présents dans le milieu cellulaire interagissent avec les groupes phosphate négatifs et contribuent à réduire la répulsion entre les phosphates et à favoriser le rapprochement des brins ou le repliement des molécules d’ADN et d’ARN.
On distingue notamment :
- des cations monovalents (Na+, K+), qui assurent une neutralisation électrostatique partielle des charges,
- des cations divalents (Mg++, Ca++), plus efficaces car leur charge plus élevée permet de neutraliser davantage de charges négatives.
2. Dans le milieu intracellulaire ou en solution expérimentale, les ions métalliques circulent librement dans l’eau mais sont entourés d’une couronne de molécules d’eau appelée sphère d’hydratation.
- Par exemple, Mg++ est généralement coordonné à six molécules d’eau dans une géométrie octaédrique.
- Ces interactions sont dynamiques et les ions peuvent se fixer temporairement puis se dissocier, contribuant ainsi à l’équilibre structural des acides nucléiques.
- Ces complexes hydratés peuvent interagir indirectement avec les acides nucléiques, notamment par des ponts hydrogène médiés par l’eau entre les molécules d’eau coordonnées et les oxygènes des groupes phosphate.
3. Dans certaines structures d’ARN complexes ou dans certains complexes protéine-ADN, des ions métalliques peuvent être coordonnés plus directement à des atomes particuliers (oxygènes des phosphates ou atomes des bases).
a. C’est notamment le cas dans :
- les ribozymes, i.e. ribozymes du groupe I ou ribozyme hammerhead, où des ions Mg++ participent à la stabilisation de la structure et parfois à la catalyse,
- les ARN ribosomiques (ARNr), dans lesquels plusieurs ions Mg++ stabilisent l’architecture du ribosome, c
- certaines structures d’ARN régulateurs, comme les riboswitches, où des ions métalliques peuvent contribuer à la stabilisation locale de la structure.
b. Dans ces situations, les ions jouent un rôle structural ou catalytique, contribuant à la stabilisation locale de la structure ou à l’activité enzymatique de certains ARN.
Agencement des bases dans l'ARN
L’ARN est généralement monocaténaire, i.e. à simple brin, mais il peut former localement des structures secondaires et tertiaires par appariements internes dans lesquels :
- l'adénine (A) s’apparie avec l'uracile (U), équivalent de la thymine dans l'ARNs, par deux liaisons hydrogène, A=U,
- la cytosine (C) s’apparie avec la guanine (G) par trois liaisons hydrogène, G≡C.
1. Mais, contrairement à l’ADN, l’ARN présente souvent :
- des appariements non canoniques, comme G•U wobble,
- des mismatches contrôlés,
- des interactions plus complexes, comme des triplets de bases ou des structures quadruplexes.
2. Cette diversité d’appariements permet la formation de nombreuses structures secondaires et tertiaires, telles que les tiges-boucles, les jonctions d’hélices ou les pseudonœuds ( structure de l'ARN).
Synthèse des interactions stabilisatrices
Les différentes interactions physicochimiques intervenant dans les acides nucléiques contribuent ensemble à la stabilité et à l’organisation structurale de l’ADN et de l’ARN. Les principales sont résumées dans le tableau suivant.
| Type d'interaction |
Mécanisme | Rôle dans l'ADN |
Rôle dans l'ARN |
|---|---|---|---|
| Liaisons hydrogène | Appariement entre bases complémentaires (A=T, G≡C) |
|
|
| Interactions π–π | Empilement des cycles aromatiques (nteractions de van der Waals entre bases) |
|
|
| Effet hydrophobe | Exclusion relative des bases aromatiques du solvant aqueux |
|
|
| Interactions ioniques | Neutralisation des charges -PO4-- par des cations (Na+, K+, Na+, Mg++…) |
Stabilisation électrostatique globale de l’ADN |
Essentielles pour la stabilisation des structure tertiaires complexes ribozymes, ARNr,…) |
| Ponts hydrogène non canoniques | Liaisons atypiques (G–U wobble, triplets…) |
Rares dans l’ADN double brin |
Très fréquents Contribution à la diversité structurale des ARN |
Retour aux bases azotées
BiochimieChimie organiqueBioénergétiqueAcides nucléiquesNucléotidesBases azotéesPentosesDifférents nucléosidesDifférents nucléotidesBiosynthèse des nucléotidesVoies des sauvetageCatabolisme des nucléotidesADNARNChromatineNucléosomesHistonesChromosomesProtidesGlucidesLipidesEnzymesCoenzymesVitaminesHormonesComposés inorganiquesTransport membranaireMoteurs moléculairesVoies de signalisation