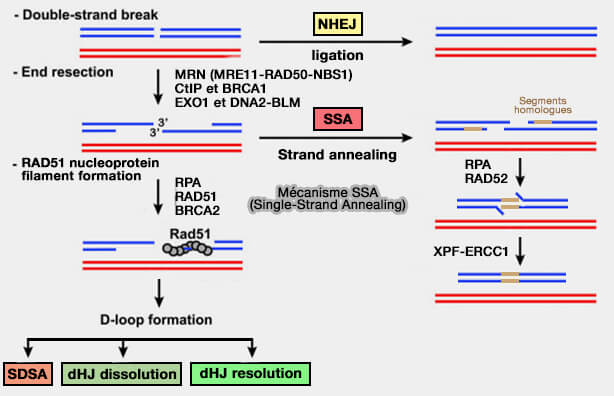
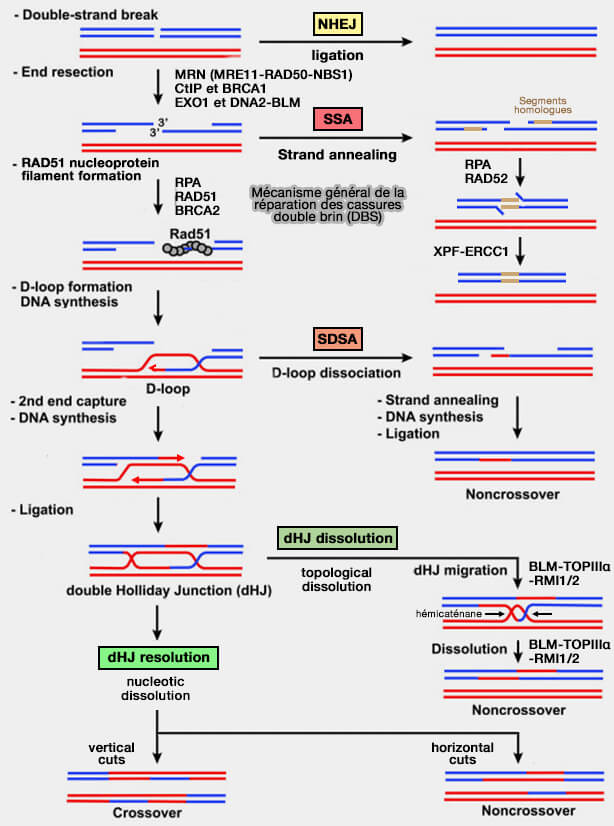
La réparation des cassures double brin peut aussi reposer sur la jonction directe des extrémités d’ADN, principalement via NHEJ, mais également par les voies mutagènes SSA et MMEJ utilisant des homologies courtes.
1. La réparation de l’ADN correspond spécifiquement à la correction enzymatique des lésions affectant la molécule d’ADN et repose sur plusieurs systèmes spécialisés :
a. réparant des lésions de l’ADN sans cassure double brin :
2. La jonction des extrémités d’ADN constitue la stratégie la plus directe de réparation des cassures double brin.
a. Dans la voie principale NHEJ, les extrémités sont reconnues et protégées par Ku70/Ku80 puis rapprochées par DNA-PKcs avant leur ligature par le complexe LIG4-XRCC4-XLF, sans nécessiter d’homologie de séquence entre les extrémités.
b. Des voies alternatives peuvent intervenir lorsque les extrémités ont subi une résection, permettant l’utilisation de courtes régions homologues (microhomologies) pour réaligner les brins avant la synthèse et la ligature.
Le mécanisme de la voie SSA intervient lorsque la résection expose des répétitions homologues longues situées de part et d’autre de la cassure.
Le mécanisme MMEJ/TMEJ s'effectue lors de l’utilisation de microhomologies courtes, généralement de 2 à 6 nucléotides, révélées après résection des extrémités d’ADN.
Ces deux mécanismes sont intrinsèquement mutagènes car l’alignement des extrémités d’ADN s’accompagne d’une perte de séquence lors du réalignement des régions homologues.
Mécanisme global des cassures double brin de l’ADN (DSB, Double-Strand Breaks)
(Figure : vetopsy.fr d'après Daley et coll)
a. Contrairement à la recombinaison homologue (HR), cette voie ne nécessite pas de séquence matrice et repose sur la reconnexion directe des extrémités d’ADN.
Ce mécanisme est actif pendant la majeure partie du cycle cellulaire, en particulier en phase G1 et G2, lorsque la chromatide sœurs n’est pas nécessairement disponible pour servir de matrice.
La voie NHEJ est rapide mais potentiellement mutagène, contrairement à la recombinaison homologue qui restaure fidèlement l’information génétique.
b.. La protéine 53BP1 contribue à orienter la réparation des cassures double brin vers le NHEJ en limitant la résection des extrémités d’ADN, notamment par le recrutement de facteurs tels que RIF1 et le complexe Shieldin, qui stabilisent les extrémités cassées et empêchent l’accès des nucléases impliquées dans la recombinaison homologue (contrôle de la résection et choix de la voie de réparation).
L’activation de BRCA1 favorise la résection des extrémités d’ADN au niveau des cassures double brin et oriente ainsi la réparation vers les mécanismes de recombinaison homologue (HR), en s’opposant à l’action de 53BP1.
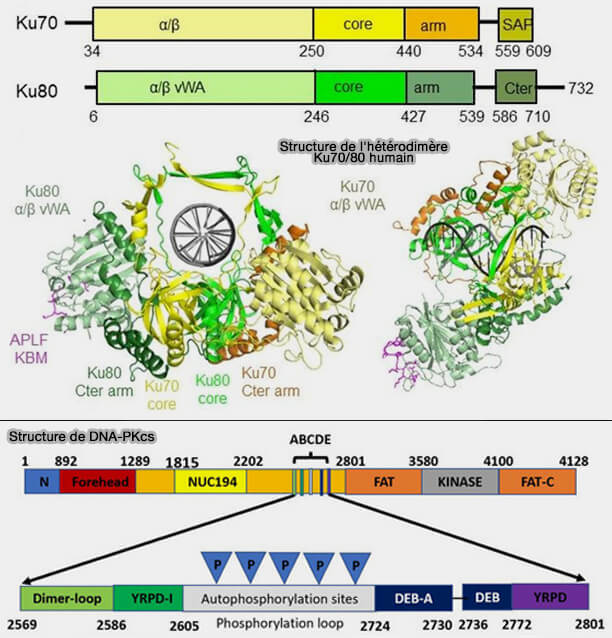
a. Ku70-Ku80 (XRCC6-XRCC5) forme un hétérodimère en anneau qui s’enfile sur l’ADN double brin au niveau de la cassure Double‐strand DNA break repair: molecular mechanisms and therapeutic targets.
Cette structure en canal central d’environ 30 Å entoure le duplex et permet au complexe de glisser légèrement le long de l’ADN tout en restant topologiquement verrouillé sur la molécule, ce qui stabilise les extrémités cassées et empêche leur dissociation ou leur résection avant l’assemblage du complexe de réparation.
Structure de l'hétérodimère Ku70/Ku80 et de DNA-PKcs
(Figure : vetopsy.fr d'après Zahid et coll et Tan et coll)
L’activation et l’autophosphorylation de DNA-PKcs provoquent un réarrangement du complexe permettant l’ajustement des extrémités d’ADN et le recrutement des facteurs responsables du traitement et de la ligation.
L'ensemble forme le complexe DNA-PK (DNA-dependent protein kinase) qui constitue la plateforme centrale du NHEJ.
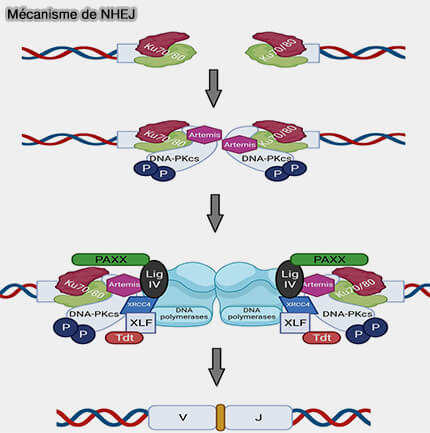
3. DNA-PKcs favorise la formation d’un complexe synaptique stabilisant la cassure en :
Mécanisme de NHEJ
(Figure : vetopsy.fr d'après Valikhani et coll)
rapprochant les extrémités d’ADN,
s’autophosphorylant, provoque un changement conformationnel du complexe DNA-PK qui agit comme une porte moléculaire et qui relâche partiellement l’ADN maintenu par Ku et permet l’accès des enzymes de traitement, notamment Artemis, aux extrémités de la cassure,
phosphorylant plusieurs protéines du NHEJ, notamment Artemis, XRCC4, XLF et les sous-unités Ku, ce qui contribue à réguler la transition vers les étapes de traitement et de ligation des extrémités d’ADN
XRCC4 et XLF peuvent également s’assembler en filaments protéiques alternés le long de l’ADN, formant une structure capable de rapprocher et de maintenir les extrémités cassées dans une configuration compatible avec la ligation par la DNA ligase IV.
4. Après la reconnaissance de la cassure, les extrémités d’ADN sont alignées et stabilisées afin de permettre leur jonction.
a. Dans de nombreux cas, les extrémités produites par la cassure ne sont pas directement compatibles pour une ligation car elles peuvent présenter des extrémités cohésives incompatibles, des surplombs simple brin, ou des bases endommagées.
PNKP (Polynucleotide Kinase Phosphatase), qui restaure les extrémités chimiques compatibles avec la ligation en générant un 5′-phosphate et un 3′-OH.
c. Des polymérases spécialisées, notamment Pol μ (POLM) et Pol λ (POLL) peuvent également intervenir pour stabiliser l’alignement des extrémités d’ADN et ajouter un nombre limité de nucléotides afin de combler de petits gaps ou d’exploiter de très courtes micro-homologies avant la ligation (Pol X DNA polymerases contribute to NHEJ flexibility 2023).
5. Une fois les extrémités rendues compatibles, la réparation se termine par la ligation des deux fragments d’ADN, rétablissant la continuité du duplex.
La ligation finale est assurée par le complexe formé par l'ADN ligase IV (LIG4), XRCC4 et XLF, qui catalyse la formation de la liaison phosphodiester finale et rétablit la continuité du duplex d’ADN.
SSA (Single-Strand Annealing)
Le mécanisme SSA (Single-Strand Annealing) est une voie de réparation des cassures double brin qui repose sur la présence de séquences répétées homologues situées de part et d’autre de la cassure.
Contrairement à la recombinaison homologue canonique (HR), ce mécanisme n’implique pas l’invasion de brin ni la formation de filaments RAD51, mais repose sur l’appariement direct de régions homologues exposées après résection de l’ADN.
2. Les régions simple brin exposées sont d’abord recouvertes par RPA, qui empêche l’appariement non spécifique des séquences et maintient l’ADN dans une conformation accessible aux protéines impliquées dans la recombinaison, notamment RAD52.
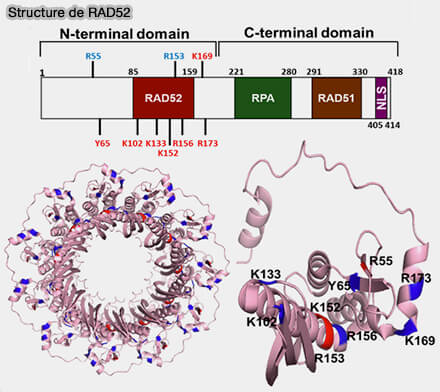
a. RAD52 s’assemble en oligomères formant des structures en anneau capables de lier simultanément plusieurs segments d’ADN simple brin.
Cette organisation permet à la protéine de capturer deux régions homologues situées de part et d’autre de la cassure et de les maintenir à proximité.
Une fois ces brins rapprochés, les bases complémentaires peuvent s’apparier directement selon les règles de Watson-Crick, ce qui conduit à la formation d’une région d’ADN double brin stabilisée par RAD52.
b. Ce mécanisme d’annealing se distingue de la recombinaison homologue médiée par RAD51 car il ne nécessite ni filament nucléoprotéique ni invasion de brin dans une molécule d’ADN double brin.
4. Après l’appariement des séquences répétées homologues, les régions situées entre la cassure et les répétitions ne peuvent pas s’apparier et restent sous forme d’ADN simple brin.
a. Ces segments excédentaires forment des structures en court volet simple brin (flap 3') qui s’étendent de part et d’autre de la région d’ADN double brin nouvellement formée par l’appariement des séquences homologues.
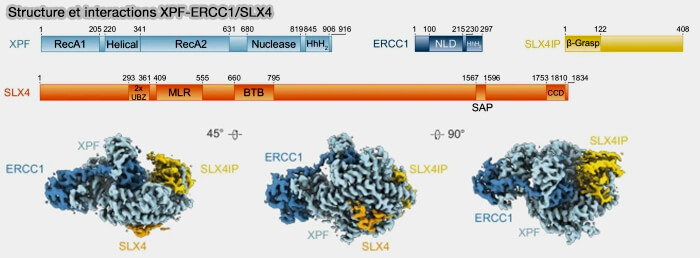
XPF-ERCC1 coupe précisément à la base du flap, au niveau de la jonction entre l’ADN double brin formé par l’appariement et le segment simple brin excédentaire.
Cette incision élimine les portions d’ADN non appariées et restaure des extrémités compatibles avec la synthèse d’ADN et la ligation ultérieure, permettant ainsi la stabilisation définitive de la molécule réparée.
c. Dans certains contextes, la nucléase XPF-ERCC1 peut être recrutée par la protéine d’échafaudage SLX4, qui agit comme une plateforme de réparation coordonnant plusieurs nucléases structure-spécifiques, notamment SLX1 et MUS81-EME1.
5. Après l’élimination des segments simple brin excédentaires, les discontinuités restantes au niveau du squelette phosphodiester sont comblées par une ADN polymérase qui complète les quelques nucléotides manquants, puis la réparation est finalisée par la ligation de l’ADN, généralement réalisée par DNA ligase I, rétablissant la continuité du duplex.
Le mécanisme SSA fusionne les deux séquences répétées homologues et supprime le segment d’ADN situé entre elles, de sorte qu’une seule copie de la répétition est conservée dans la molécule réparée.
MMEJ (Microhomology-Mediated End Joining)
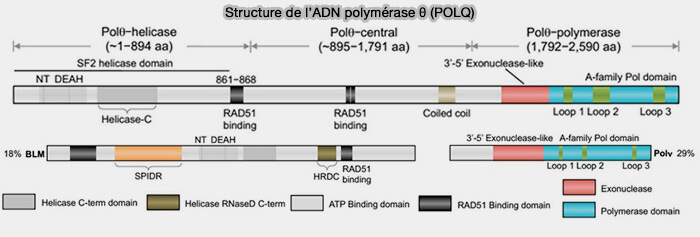
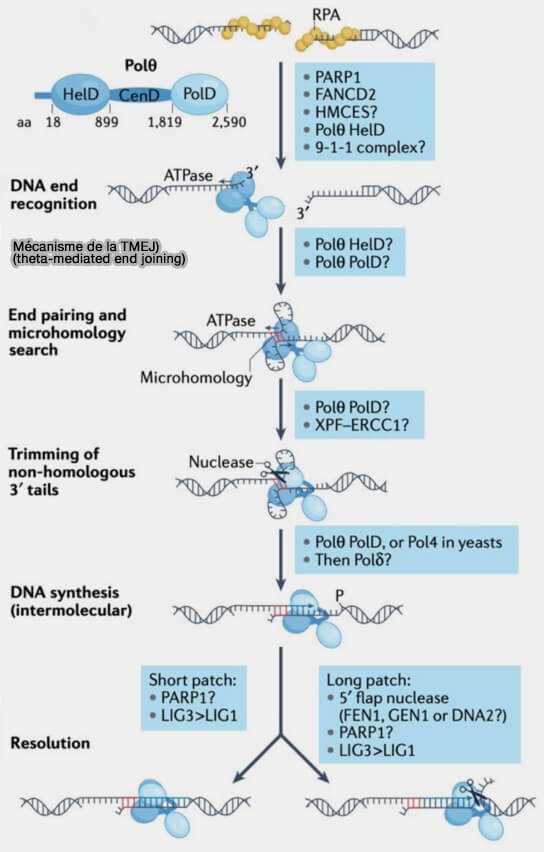
La microhomology-mediated end joining (MMEJ), également appelée theta-mediated end joining (TMEJ), constitue une voie de réparation des cassures double brin qui repose sur l’utilisation de courtes microhomologies et dépend de l’activité de l’ADN polymérase θ (POLQ).
la résection peut être étendue par l'exonucléase EXO1 ou par le système BLM-DNA2, ce qui génère des extrémités simple brin 3′.
2. La divergence entre les deux mécanismes apparaît lors de l’étape d’appariement des séquences homologues.
a. Dans la SSA, l’alignement s’effectue entre de longues séquences répétées, souvent séparées par plusieurs centaines ou milliers de nucléotides, ce qui conduit à la formation d’un duplex étendu stabilisé par RAD52 et suffisamment stable pour se maintenir sans polymérase spécialisée.
b. À l’inverse, la MMEJ exploite uniquement de courtes microhomologies, généralement longues de 2 à 20 nucléotides, présentes à proximité de la cassure.
L’appariement est intrinsèquement instable car deux extrémités résectées qui ne partagent que quelques nucléotides complémentaires ne restent généralement pas alignées spontanément.
Il faut donc un facteur capable de stabiliser cet appariement transitoire.
un domaine central impliqué notamment dans l’interaction avec RAD51,
un domaine polymérase de type A situé à l’extrémité C-terminale.
b. Le domaine polymérase de Pol θ est très atypique car il présente :
une poche catalytique plus ouverte,
des insertions spécifiques (loops) absentes des polymérases réplicatives,
une tolérance élevée aux mésappariements et aux duplex très courts
Structure de l’ADN polymérase θ (POLQ)
(Figure : vetopsy.fr d'après Black et coll)
b. Dans la figure, BLM est représentée comme exemple d’hélicase de réparation de l’ADN impliquée notamment dans la recombinaison homologue (HR) et la dissolution des jonctions de Holliday, tandis que Pol ν est montrée comme exemple de polymérase humaine de la famille A impliquée dans certains mécanismes de réparation et de tolérance aux lésions de l’ADN.
déplaçant ou en réorganisant les complexes RPA qui recouvrent l’ADN simple brin pour le stabiliser mais empêchent également l’appariement spontané des microhomologies,
rapprochant les extrémités résectées,
favorisant l’exposition de microhomologies compatibles.
Remarque : le domaine hélicase agit donc davantage comme un facteur de rapprochement et de stabilisation des extrémités d’ADN que comme une hélicase processive classique.
Mécanisme de la TMJE (MMEJ)
(Figure : vetopsy.fr d'après Xan)
5. Lorsque les microhomologies s’apparient, les extrémités d’ADN sont rapprochées, mais des segments simple brin non appariés subsistent de part et d’autre de la jonction, correspondant aux séquences situées entre la cassure initiale et les microhomologies utilisées pour l’alignement.
Ces segments excédentaires sont ensuite éliminés par des endonucléases capables de cliver les jonctions ADN simple brin/double brin, notamment l'endonucléase XPF-ERCC1, comme dans la SSA, qui incise les régions non appariées.
6. Une fois les microhomologies exposées, son domaine polymérase de type A situé à l’extrémité C-terminale, contrairement aux polymérases réplicatives classiques, agit comme un facteur de stabilisation de la jonction et peut :
accepter et aligner des structures d’ADN imparfaitement appariées,
initier la synthèse à partir de microhomologies très courtes, de 2–6 nucléotides, ce qui permet d’étendre les extrémités 3′ appariées et de consolider l’intermédiaire de réparation,
poursuivre la synthèse après un réalignement du brin naissant.
Remarque : dans certains cas, la polymérase θ peut décrocher de la matrice et se réaligner sur une autre microhomologie proche, un phénomène appelé template switching, qui peut générer de courtes insertions ou des duplications locales au point de jonction caractéristiques de la voie TMEJ (Template switching in DNA replication can create and maintain RNA hairpins 2022).
Ces signatures mutationnelles associant microhomologies, délétions et petites insertions sont fréquemment observées dans les génomes tumoraux présentant une activité élevée de la voie MMEJ.
Le mécanisme MMEJ, comme SSA, fusionne les deux séquences homologues utilisées pour l’appariement et supprime le segment d’ADN situé entre elles, de sorte qu’une seule copie de la microhomologie est conservée dans la molécule réparée.




 contrôle de la résection et choix de la voie de réparation).
contrôle de la résection et choix de la voie de réparation).