


Acides nucléiques
Chromatine
Histones : types et variants
- Biochimie
- Chimie organique
- Bioénergétique
- Composition de la matière vivante
- Glucides
- Lipides
- Coenzymes
- Hormones
- Composés inorganiques

Les histones se répartissent en histones du cœur (H2A, H2B, H3 et H4) formant l’octamère du nucléosome et en histones de liaison H1 qui stabilisent l’ADN linker et participent à l’organisation de la chromatine.
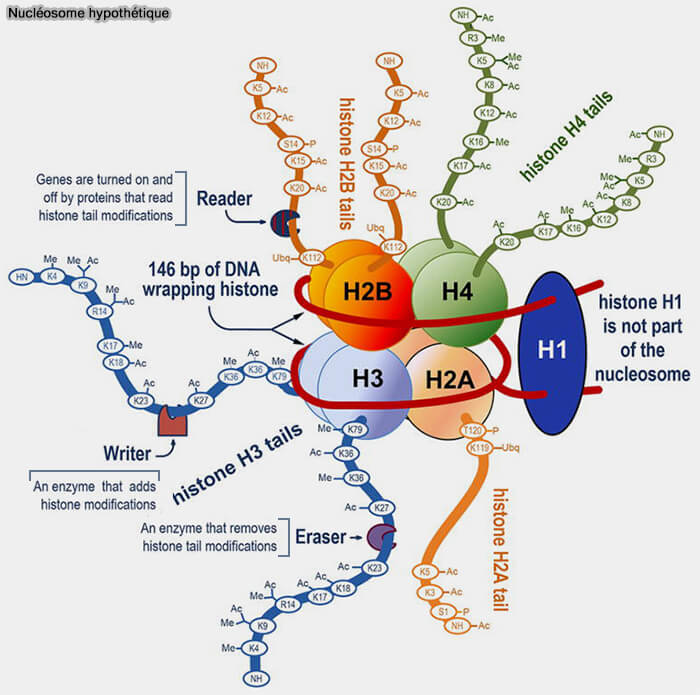
(Figure : vetopsy.fr d'après Régnier et Kim)
H2A, H2B, H3 et H4 : octomère central du nucléosome
Les histones H2A, H2B, H3 et H4 s’assemblent pour former l’octamère histonique autour duquel s’enroule l’ADN pour former le nucléosome ( Interactions entre DNA et histones de coeur).
Interactions entre DNA et histones de coeur).
Contrairement à l’histone H1, qui se fixe sur l’ADN de liaison, ces histones constituent les protéines du cœur du nucléosome.
Ce sont des petites protéines d’environ 102 à 135 acides aminés, très riches en acides aminés basiques (lysine et arginine), ce qui favorise les interactions électrostatiques avec le squelette phosphodiester de l’ADN.
Elles figurent parmi les protéines les plus conservées chez les eucaryotes, certaines histones comme H4 ne présentant que quelques différences d’acides aminés entre des organismes très éloignés phylogénétiquement.
H3 et H4
Le cœur structural de l’octamère est formé par les dimères d'histones H3 et H4 (dimérisation et assemblage des histones).
H3
H3 est l’une des histones centrales du nucléosome et joue un rôle majeur dans l’intégrité structurale de l’octamère ainsi que dans la régulation fonctionnelle de la chromatine.
Elle s’associe avec l’histone H4 pour former le dimère H3-H4, puis le tétramère (H3-H4)2 qui constitue le cœur de l’octamère histonique.
L’histone H3 comprend plusieurs variants qui diffèrent par leur mode d’incorporation et leur localisation dans la chromatine ( variants d'histones).
1. H3.1 et H3.2 sont des histones dites de réplication, incorporées dans la chromatine lors de la réplication de l’ADN et déposées principalement par le chaperon d’histones CAF-1.
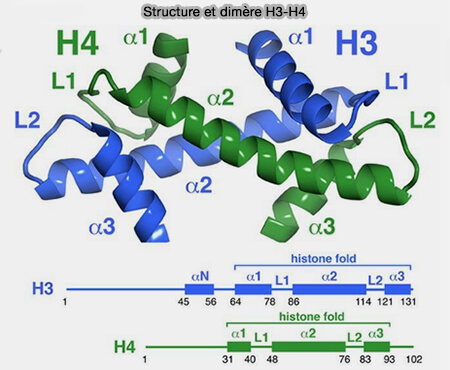
(Figure : vetopsy.fr d'après MacGinty et Tan)
2. H3.3 est une variante indépendante de la réplication, associée aux régions de chromatine active et impliquée dans la transcription des gènes (euchromatine).
Son incorporation est assurée par des chaperons d’histones, notamment :
- le complexe HIRA qui dirige son dépôt dans les gènes actifs et les régions transcrites,
- le complexe DAXX-ATRX qui l’incorpore dans certaines régions répétées du génome, notamment les télomères et l'hétérochromatine péricentromérique.
3. CENP-A constitue une variante spécialisée de H3 localisée au niveau des centromères, où elle remplace l’histone H3 canonique dans les nucléosomes centromériques et participe à l’organisation du centromère et à l’assemblage du kinétochore.

Le centromère et les protéines CENP (CENtromere Proteins) sont étudiés dans des pages spécifiques.
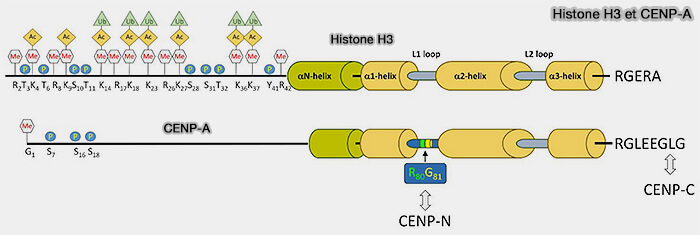
(Figure : vetopsy.fr d'après Sharma et coll)
4. Les extrémités N-terminales de l’histone H3 sont le siège de nombreuses modifications post-traductionnelles qui participent à la régulation de l’organisation et de l’activité de la chromatine ( code des histones).
a. Ces modifications comprennent notamment des méthylations de résidus lysine (K4, K9, K27, K36), ainsi que des acétylations, phosphorylations et ubiquitinations, qui modulent l’accessibilité de la chromatine et l’activité transcriptionnelle.
b. Certaines marques de H3 sont particulièrement caractéristiques de certains états chromatiniens :
- H3K4me3 est associée aux promoteurs actifs,
- H3K36me3 à l’élongation transcriptionnelle dans les régions codantes des gènes actifs,
- H3K9me3 à l’hétérochromatine constitutive,
- H3K27me3 aux domaines réprimés dépendant des complexes Polycomb,
- H3S10ph, phosphorylation de la sérine 10 de l’histone H3 (H3S10ph), notamment par la kinase Aurora B, est associée à la condensation des chromosomes mitotiques.
c. Ces marques jouent un rôle clé dans :
- la transcription (activation ou répression),
- l’organisation des domaines de chromatine tels que l’euchromatine et l’hétérochromatine,
- certains mécanismes épigénétiques comme le bookmarking mitotique et la transmission de la mémoire épigénétique.
H4
H4 est l’une des histones les plus conservées chez les eucaryotes.
Contrairement aux histones H3 et H2A, qui possèdent plusieurs variants fonctionnels, l’histone H4 ne présente pratiquement pas de variants spécialisés ( variants d'histones).
1. H4 contribue à la stabilité de l’octamère histonique par ses interactions avec :
- H3, le noyau central du nucléosome en formant le tétramère (H3-H4)2,
- les dimères H2A-H2B, par l'interaction de sa queue N-terminale,
- l’ADN enroulé autour du cœur nucléosomique.
2. La queue N-terminale de H4 sort du nucléosome et peut interagir avec le patch acide porté par le dimère H2A-H2B d’un nucléosome voisin, établissant des interactions internucléosomiques qui rapprochent les nucléosomes et contribuent à la compaction de la chromatine pour l’organisation de structures chromatiniennes de niveau supérieur (The human telomeric nucleosome displays distinct structural and dynamic properties 2020).
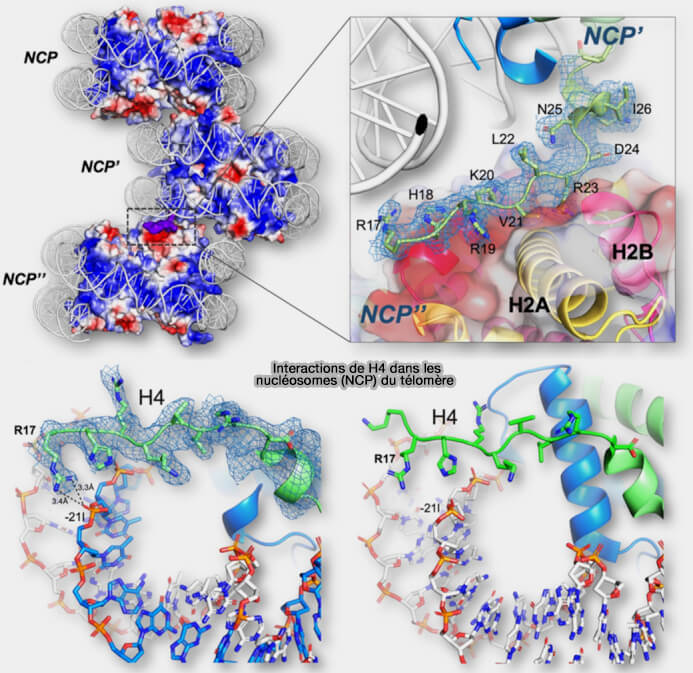
(Figure : vetopsy.fr d'après Soman et coll)
a. Les modifications post-traductionnelles de H4, notamment l’acétylation et la méthylation de résidus lysine de la queue N-terminale ( code des histones) :
- modulent ces interactions entre nucléosomes et contribuent à la compaction de la chromatine et à l’organisation de structures chromatiniennes de niveau supérieur,
- influencent l’activité transcriptionnelle (Effect of histone H4 tail on nucleosome stability and internucleosomal interactions 2021).
b. Un exemple particulièrement bien caractérisé est l’acétylation de la lysine 16 de H4 (H4K16ac).
- Cette modification neutralise la charge positive de la lysine, ce qui réduit l’interaction de la queue N-terminale de H4 avec le patch acide H2A-H2B d’un nucléosome voisin.
- La diminution de ces interactions internucléosomiques limite la compaction de la chromatine et favorise une organisation plus ouverte compatible avec l’activité transcriptionnelle.
Remarque : R19, K20 et R23 sont aussi impliqués dans cette interaction.
Si les modifications de H3 écrivent l’état chromatinien (activation ou répression des gènes), celles de H4 règlent surtout l’architecture physique de la chromatine en modulant sa compaction, notamment entre euchromatine et hétérochromatine.
H2A et H2B
Organisation structurale du dimère H2A-H2B
Les histones H2A et H2B forment des dimères H2A-H2B qui viennent se fixer de part et d’autre du tétramère (H3-H4)2, complétant l’assemblage de l’octamère histonique du nucléosome.
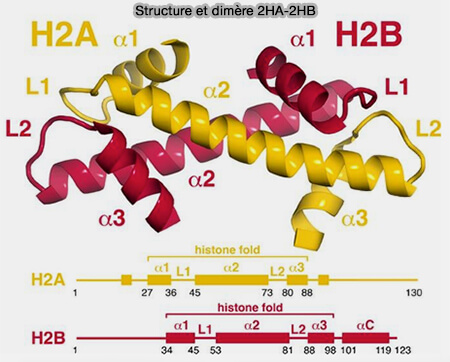
(Figure : vetopsy.fr d'après MacGinty et Tan)
Les H2 stabilisent l’ADN enroulé autour du nucléosome et participent à la formation de la fibre de 10 nm.
Le dimère H2A-H2B constitue la partie la plus dynamique du nucléosome.
Contrairement au tétramère central (H3-H4)2, qui est très stable, les dimères H2A-H2B peuvent être retirés et réincorporés par des chaperons d’histones et des complexes de remodelage de la chromatine.
Cette dynamique permet l’échange de variants d’histones (par exemple H2A.Z, H2A.X ou macroH2A) et contribue à la plasticité structurale et fonctionnelle des nucléosomes lors de la transcription, de la réplication ou de la réparation de l'ADN.
Patch (tache) acide du nucléosome (acidic patch)
1. Le patch (tache) acide H2A-H2B (acidic patch) est une région de surface fortement chargée négativement située sur le dimère d’histones H2A-H2B à la surface du nucléosome (Interactions of Nucleosomes with Acidic Patch-Binding Peptides: A Combined Structural Bioinformatics, Molecular Modeling, Fluorescence Polarization, and Single-Molecule FRET Study 2023).
Il est formé principalement par des résidus acides de H2A, notamment des glutamates (E) et des aspartates (D), ainsi que par quelques résidus acides de H2B, et constitue l’un des principaux sites d’interaction du nucléosome avec d’autres protéines de la chromatine (The role of the nucleosome acidic patch in modulating higher order chromatin structure 2013).
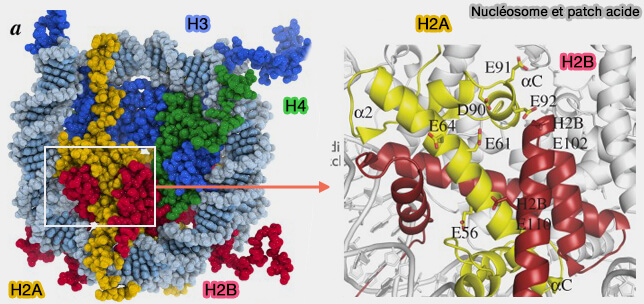
(Figure : vetopsy.fr modifiée d'après MacGinty et Tan et Kalashnikova et coll)
2. Le patch acide constitue une plateforme d’interaction pour de nombreuses protéines régulatrices de la chromatine.
a. Certaines protéines, comme CENP-C, LANA, RAP1 ou la sous-unité Spt16 du complexe FACT, interagissent avec cette région en utilisant des résidus basiques qui s’insèrent dans une poche de la surface acide du nucléosome.
Ces interactions reposent fréquemment sur un mécanisme appelé " arginine anchor ", dans lequel un résidu arginine de la protéine partenaire s’insère dans la cavité formée par les résidus acides du dimère H2A-H2B.
b. Le patch acide joue un rôle fonctionnel majeur en servant de site d’interaction pour la queue N-terminale de l’histone H4 d’un nucléosome voisin, contribuant ainsi aux interactions internucléosomiques impliquées dans l’organisation de la chromatine.
Variants et modifications post-traductionnelles
1. L’histone H2A possède plusieurs variants spécialisés ( variants d'histones).
- H2AX, environ 10 % des histones H2A chez les mammifères, intervient dans la réponse aux cassures double brin de l’ADN lors de sa phosphorylation pour former γH2AX (H2AXS139ph).
- H2AZ est enrichie au niveau des promoteurs et participe à la régulation de la transcription ainsi qu’à la plasticité de la chromatine.
- macroH2A est associée à l’hétérochromatine facultative et contribue notamment à l’inactivation du chromosome X.
Les modifications post-traductionnelles de H2A comprennent notamment l’acétylation, la méthylation, la phosphorylation et l’ubiquitination, qui participent à la régulation de ( code des histones) :
- la transcription, par exemple l’acétylation de H2A aux promoteurs actifs,
- la réparation de l’ADN, par exemple la phosphorylation de H2AX en γH2AX lors des cassures double brin qui est une marque clé du recrutement des complexes de réparation,
- l’organisation de la chromatine, par exemple l’ubiquitination de H2AK119 associée à la répression dépendante des complexes Polycomb.
2. Contrairement à l’histone H2A, qui possède plusieurs variants fonctionnels importants, l’histone H2B présente relativement peu de variants ( variants d'histones).
La régulation fonctionnelle de H2B repose surtout sur certaines modifications post-traductionnelles, en particulier l’ubiquitination spécifique sur la lysine 120 (H2BK120ub1).
Déposée lors du passage de l’ARN polymérase II sur les gènes transcrits, cette modification favorise l’élongation transcriptionnelle et stimule la méthylation de certaines lysines de l’histone H3, notamment H3K4 et H3K79, illustrant un mécanisme de coopération entre modifications d’histones.
Histone H1 : histone de liaison
Structure et variants de H1
1. L'histone H1, appelée histone de liaison ou histone linker, se fixe sur l’ADN de liaison reliant deux nucléosomes adjacents, stabilise l’organisation de la fibre de chromatine et favorise ainsi la formation de structures chromatiniennes de niveau supérieur.
a. Contrairement aux histones du cœur du nucléosome (H2A, H2B, H3 et H4), H1 :
- ne fait pas partie de l’octamère histonique,
- ne présente pas d’homologie structurale avec ces histones,
- est présente en une seule molécule par nucléosome, dans un rapport stoechiométrique proche de 1:1.
b. L’association de l’histone H1 avec le nucléosome et l’ADN de liaison forme une particule appelée chromatosome.

Vous pouvez lire : The missing linker: emerging trends for H1 variant-specific functions (2021) et Distinct Structures and Dynamics of Chromatosomes with Different Human Linker Histone Isoforms (2021) avec des vidéos.
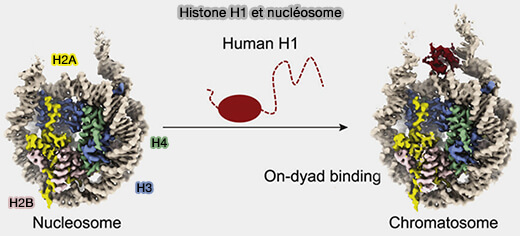
(Figure : vetopsy.fr d'après Zhou et coll)
1. H1 se fixe au point d’entrée et de sortie de l’ADN sur le nucléosome, au niveau central de la dyade stabilisant l’enroulement autour de l’octamère.
a. Sa structure, plus volumineuse que les histones du coeur du nucléosome et non encore résolue, comprend :
- une queue N-terminale courte et flexible, impliquée dans les interactions avec l’ADN et certaines protéines de la chromatine,
- un domaine central globulaire, en forme d'aile, responsable de la liaison spécifique au nucléosome,
- une longue queue C-terminale très basique, qui joue un rôle majeur dans la stabilisation des fibres chromatiniennes et la compaction de la chromatine.
b. Une grande partie des queues N- et C-terminales sont intrinsèquement désordonnées, ce qui rend difficile la résolution de la structure complète de l’histone H1 associée au nucléosome.
Très récemment, à l’aide de techniques de cryo-EM de pointe, la structure de H1 (H5) aviaire liée à un dédocanucléosome a été déterminée à une résolution de 3,6 Å, révélant la structure atomique d’une histone H5 presque pleine longueur (Distinct Structures and Dynamics of Chromatosomes with Different Human Linker Histone Isoforms 2021 avec des vidéos).
(Figure : vetopsy.fr d'après Zhou et coll)
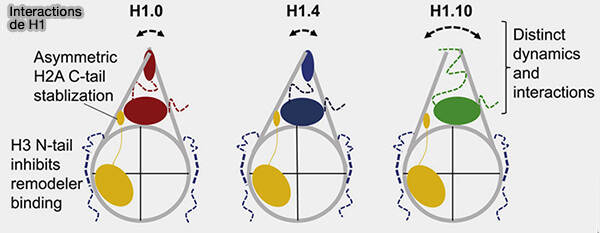
2. L’histone H1 existe sous plusieurs variants, H1.1 à H1.X ( variants d'histones).
a. Chez l’homme, on en dénombre onze, dont sept sont exprimés dans les cellules somatiques et quatre principalement dans les cellules germinales,.
b. Certains variants présentent des propriétés structurales ou fonctionnelles spécifiques, contribuant à la diversité des états chromatiniens.
- Par exemple, H1.0 (H1°) est associé aux cellules différenciées et à une chromatine plus condensée et peu active transcriptionnellement.
- H1.X est souvent enrichie dans des régions transcriptionnellement actives.
- Certains variants germinaux comme H1t participent à l’organisation particulière de la chromatine lors de la spermatogenèse (La compaction de la chromatine au cours de la spermatogenèse : Rôle des bromodomaines de la protéine Brdt 2009).
Fonctions de H1
Les principales fonctions de l’histone H1 sont liées à l’organisation et à la régulation de la chromatine.
1. H1 participe à la compaction de la chromatine en se fixant au niveau de la dyade du nucléosome et en stabilisant l’ADN entrant et sortant, ce qui rapproche les nucléosomes voisins et favorise l’organisation de fibres chromatiniennes plus compactes.
En stabilisant l’ADN entrant et sortant du nucléosome, H1 :
- limite la mobilité des nucléosomes le long de l’ADN et contribue ainsi à la stabilisation de la structure chromatinienne.
- réduit l’accès de l’ADN aux facteurs de transcription et aux complexes de transcription.
2. H1 participe aussi aux changements d’organisation de la chromatine au cours du cycle cellulaire, notamment lors de la condensation des chromosomes mitotiques par des phosphorylations catalysées notamment par des kinases mitotiques, en particulier Aurora B, qui modulent son interaction avec la chromatine.
3. H1 est une histone dynamique dont l’association avec la chromatine varie selon l’état transcriptionnel de la cellule.
- Elle peut subir différentes modifications post-traductionnelles, notamment des acétylations et des phosphorylations, qui modulent sa capacité à condenser la chromatine ( code des histones).
- Le variant H1.0 (H1°) est moins mobile que les autres isoformes de H1 et contribue à la stabilisation d’une chromatine fortement condensée dans les cellules différenciées.
Remarque : H5 est une histone linker spécialisée, analogue à H1, présentant une affinité plus forte pour l’ADN et favorisant une compaction chromatinienne plus stable.
Elle est notamment présente dans les globules rouges aviaires, où elle contribue au silencement transcriptionnel presque complet de la chromatine.
Code des histones
BiochimieChimie organiqueBioénergétiqueAcides nucléiquesNucléotidesADNARNChromatineStructure dynamique de la chromatineNucléosomesHistonesMembres des histonesCode des histonesEuchromatine, hétérochromatine et remodelageChromosomesStructure des chromosomesCentromèreTélomèreProtidesGlucidesLipidesEnzymesCoenzymesVitaminesHormonesComposés inorganiquesTransport membranaireMoteurs moléculairesVoies de signalisation