


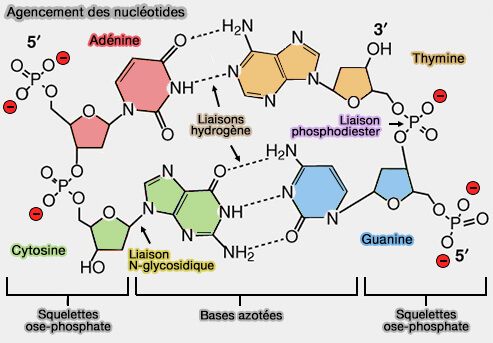
Acides nucléiques
ADN : réparation
sans cassure double brin
Réparation par excision de base (BER)
Réparation par excision de nucléotides (NER)
- Biochimie
- Chimie organique
- Bioénergétique
- Composition de la matière vivante
- Transport membranaire
- Moteurs moléculaires
- Voies de signalisation

Les réparations BER et NER assurent l’élimination des bases altérées ou des lésions volumineuses de l’ADN par excision du segment endommagé suivie d’une resynthèse du brin réparé.
La réparation de l’ADN correspond spécifiquement à la correction enzymatique des lésions affectant la molécule d’ADN et repose sur plusieurs systèmes spécialisés :
a. réparant des lésions de l’ADN sans cassure double brin :
b. impliquant des cassures double brin (DSBR) :
- HR (recombinaison homologue)
- la réparation par jonction des extrémités d’ADN (NHEJ, SSA, MMEJ/TMEJ).
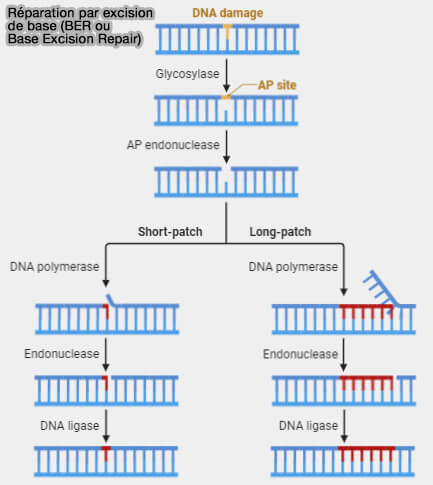
Réparation par excision de base (BER)
Principe général
La réparation par excision de base (BER ou Base Excision Repair) est un mécanisme de réparation de l’ADN qui corrige les altérations chimiques affectant une base individuelle sans perturber la structure globale de la double hélice (Base Excision Repair 2013).
Ce système intervient notamment lors de désaminations, d’oxydations ou d’alkylations des bases azotées.
1. Une ADN glycosylase spécifique reconnaît la base endommagée et hydrolyse la liaison N-glycosidique qui la relie au désoxyribose.
L’excision de la base crée un site abasique, appelé site AP (apurinique/apyrimidinique), où l'ose reste présent mais sans base azotée.
(Figure : vetopsy.fr d'après OpenStax)
{kind=link}
2. L'AP endonucléase APE1 clive la liaison phosphodiester du côté 5' du site AP, générant une extrémité 3'-OH utilisable par l’ADN polymérase pour préparer la réparation.
Cette incision génère également une extrémité 5' portant un groupement désoxyribose-phosphate (5'-dRP) qui devra être éliminé avant la synthèse complète du brin réparé.
Remarque : une enzyme apparentée, APE2 (APEX2), possède une activité endonucléase beaucoup plus faible et agit surtout comme exonucléase 3' → 5' dans certaines réponses au stress réplicatif et dans la voie de signalisation ATR activée lors de la présence de fourches de réplication endommagées.
(Figure : vetopsy.fr d'après biorender.com)
3. Une ADN polymérase insère le nucléotide correct en utilisant le brin complémentaire comme matrice (The base excision repair process: comparison between higher and lower eukaryotes 2021).
a. Dans la réparation courte (short-patch BER), la polymérase, généralement l’ADN polymérase β, remplace uniquement le nucléotide manquant au niveau du site AP.
Le groupement 5'-dRP est généralement éliminé par l’activité désoxyribose-phosphate lyase (dRP lyase) portée par l’ADN polymérase β.
b. Dans la réparation longue (long-patch BER), après l’incision du site AP, les polymérases δ ou ε associées aux facteurs PCNA et RFC ne remplacent pas uniquement le nucléotide manquant, mais poursuivent la synthèse sur plusieurs nucléotides (environ 2 à 10), en utilisant le brin complémentaire comme matrice.
- Cette synthèse pousse le segment d’ADN situé en aval, qui forme un court volet simple brin (flap) contenant les nucléotides initiaux.
- Ce volet d’ADN déplacé est ensuite éliminé par l’endonucléase FEN1 (Flap Endonuclease 1), qui clive ce segment simple brin avant la ligature finale.
4. Une ADN ligase restaure la continuité du squelette phosphodiester en reliant les extrémités du brin réparé.
- Le complexe ADN ligase III-XRCC1 assure le plus souvent la ligature dans le short-patch BER.
- L'ADN ligase I peut intervenir lors du long-patch BER associé aux facteurs de réplication.
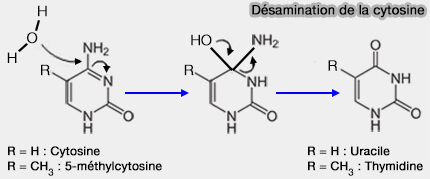
Exemple : réparation de la désamination de la cytosine en uracile
La désamination spontanée de la cytosine entraîne la transformation de la cytosine en uracile dans l’ADN, où cette base est normalement absente.
(Figure : vetopsy.fr)
Ce type de lésion est fréquent, i.e. plusieurs centaines par jour et par cellule, et représente une source majeure de mutations G≡C ➞ A=T si la lésion n’est pas réparée.
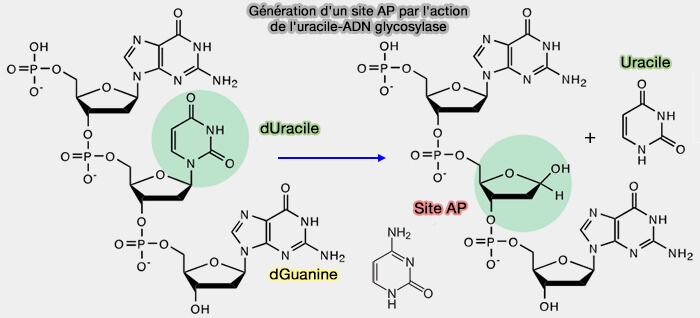
1. La présence d’uracile dans l’ADN est reconnue par l’uracile-ADN glycosylase (EC 3.2.2.27).
- L'uracile-ADN glycosylase hydrolyse la liaison N-glycosidique entre le carbone 1' du désoxyribose et l'atome N1 de l’uracile anormalement présent dans l'ADN.
- Les produits de la réaction sont de l'uracile libre et un brin d'ADN contenant un site abasique, i.e. sans base appelé site AP (apurinic/apyrimidinic site).
2. Puis, on assiste à un clivage du squelette ose-phosphate par l'AP endonucléase APE1 qui coupe l’ADN du côté 5' au niveau du site abasique.
3. L'ADN polymérase β insère une nouvelle cytosine et l’ADN ligase III associée à XRCC1 referme le brin d’ADN.
(Figure : vetopsy.fr)
Réparation par excision de nucléotides (NER)
Principe général
La réparation par excision de nucléotides (NER ou Nucleotide Excision Repair) est un mécanisme de réparation de l’ADN qui élimine des lésions volumineuses perturbant la géométrie de la double hélice (Nucleotide Excision Repair).
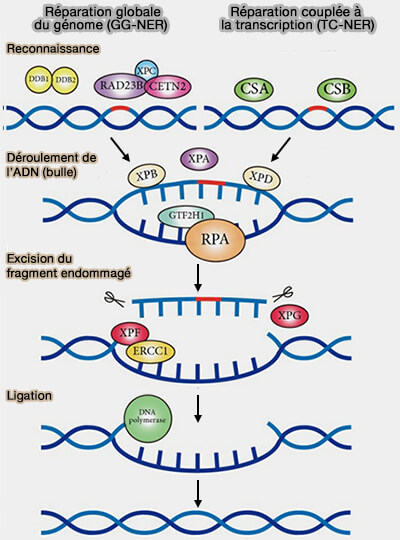
(Figure : vetopsy.fr d'après Liu et coll)
1. Ce système corrige notamment les lésions qui déforment localement la double hélice d’ADN comme :
- les dimères de pyrimidines induits par les UV, correspondant à des liaisons covalentes entre les carbones C5 et C6 de deux bases adjacentes du même brin d’ADN, ce qui rapproche les deux bases et les incline l’une vers l’autre, perturbant l’empilement normal des bases et provoquant une distorsion locale de la double hélice,
- les photoproduits 6-4, dans lesquels une liaison anormale se forme entre le carbone 6 d’une pyrimidine et le carbone 4 de la pyrimidine voisine du même brin d’ADN,
- les adduits chimiques (DNA adduct) qui correspondent à la fixation covalente d’un groupement chimique relativement volumineux sur une base de l’ADN formés par des agents mutagènes.
Des exemples typiques d’adduits de l’ADN sont les adduits benzo[a]pyrène–guanine formés par des composés de la fumée de tabac, les adduits aflatoxine–guanine produits par certaines toxines fongiques, ainsi que les adduits ADN–cisplatine induits par cet agent chimiothérapeutique.
2. Le principe du NER, qui repose sur l’excision d’un court fragment d’ADN contenant la lésion, suivie de la resynthèse du segment manquant en utilisant le brin intact comme matrice (The Differential Expression of Core Genes in Nucleotide Excision Repair Pathway Indicates Colorectal Carcinogenesis and Prognosis 2018).
3. Chez les eucaryotes, la reconnaissance des lésions dans la voie NER repose sur deux sous-voies qui se distinguent uniquement par leur mécanisme de détection initiale de la lésion, le mécanisme d’excision et de réparation étant commun :
- la réparation globale du génome (GG-NER),
- la réparation couplée à la transcription (TC-NER).
Reconnaissance des lésions
Réparation globale du génome (GG-NER)
Reconnaissance directe par le complexe XPC
La réparation globale du génome (GG-NER) détecte les lésions sur l’ensemble du génome.
La reconnaissance des déformations de la double hélice est assurée par des complexes de surveillance de l’ADN principalement constitués par le complexe XPC qui parcoure le génome en se liant transitoirement à l’ADN et en détectant les perturbations locales de la double hélice provoquées par les lésions.
1. Le complexe XPC est constitué de la protéine XPC associée à la protéine HR23B (RAD23B), qui stabilise XPC et favorise sa liaison à l’ADN endommagé.
a. Ce complexe peut également s’associer à la protéine centrin-2 (CETN2), formant l’ensemble fonctionnel XPC-HR23B-CETN2 impliqué dans la reconnaissance des lésions lors du GG-NER.
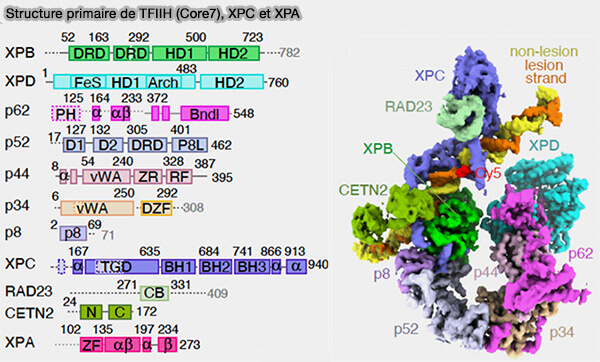
b. Le complexe XPC reconnaît la lésion et recrute ensuite le complexe TFIIH, qui contient notamment les hélicases XPB et XPD, lesquelles déroulent localement l’ADN afin de permettre la vérification et l’excision de la lésion.
- Le complexe TFIIH est le même complexe multiprotéique que celui participant à l’assemblage du complexe de pré-initiation transcriptionnelle (PIC) des gènes transcrits par l’ARN polymérase II.
- Toutefois, dans le NER, TFIIH est recruté après la reconnaissance de la lésion et intervient principalement dans l’ouverture locale de l’ADN autour du site endommagé
(Figure : vetopsy.fr d'après Kim et coll)
Remarque : Cy5 est un colorant fluorescent cyanine expérimental utilisé pour marquer l’ADN ou une protéine dans les expériences.
2. XPC comprend :
a. deux longues hélices α :
- LHN (Long Helix N-terminal) qui interagit notamment avec la sous-unité p62 du complexe TFIIH et contribue à orienter TFIIH sur l’ADN endommagé,
- LHC (Long Helix C-terminal) qui traverse la région de l’ADN et interagit avec la sous-unité XPB de TFIIH, interaction stabilisée par la protéine centrin-2 (CETN2),
b. les domaines BHD2 et BHD3 dont les β-hairpins peuvent pénètrer dans la double hélice.
2. Le complexe XPC ne reconnaît pas directement la base endommagée elle-même, mais les lésions qui rendent la double hélice moins stable thermodynamiquement, notamment :
- une distorsion locale de la double hélice,
- un désappariement local des bases,
- une perte d’empilement des bases (π-stacking),
- une tendance des bases à s’extruder légèrement hors de l’hélice.
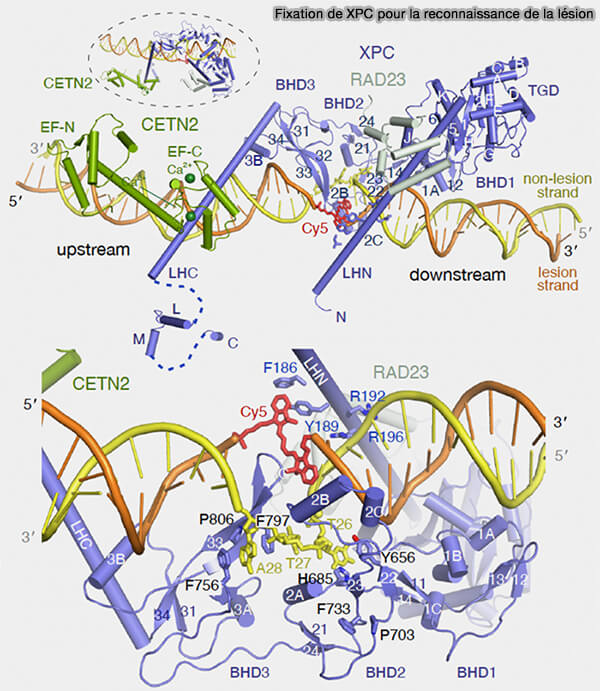
(Figure : vetopsy.fr d'après Kim et coll)
3. Le complexe XPC-HR23B-CETN2 diffuse le long de l’ADN et établit des contacts transitoires avec le squelette ose-phosphate pour tester la stabilité locale de l’hélice.
a. Lorsque l’ADN est normal, les bases bien empilées et la double hélice rigide, XPC ne se fixe pas.
b. Si une lésion provoque un désappariement local ou une perturbation de l’empilement des bases, XPC insère les β-hairpins des domaines BHD2 et BHD3 dans l’ADN.
- Cette insertion provoque une déstabilisation de l’appariement local et une ouverture locale de l’hélice en séparant les deux brins sur quelques nucléotides, facilitant la reconnaissance de la lésion et préparant la vérification de la lésion par l’hélicase XPD du complexe TFIIH.
- Elle stabilise la fixation du complexe XPC sur l’ADN endommagé, rendant cette interaction thermodynamiquement favorable.
- Elle provoque le basculement des bases du brin lésé hors de l’hélice.
Reconnaissance assistée par le complexe DDB1-DDB2
1. Dans le cas de certaines lésions induites par les UV, notamment les dimères de pyrimidines dans lesquelles la déformation est faible, la reconnaissance initiale est facilitée par le complexe DDB1-DDB2 (UV-DDB), constitué des protéines DDB1 et DDB2 (The Molecular Basis of CRL4DDB2/CSA Ubiquitin Ligase Architecture, Targeting, and Activation 2011).
-
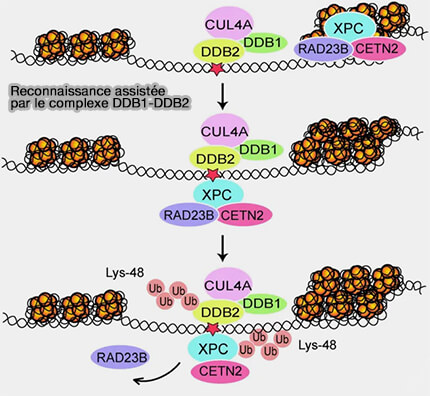
Reconnaissance assistée par le complexe DDB1-DDB2
(Figure : vetopsy.fr d'après Rüthemann et coll)
a. La protéine DDB2 possède une poche de reconnaissance capable de se lier directement à la base lésée et de la faire basculer hors de la double hélice, ce qui permet la reconnaissance initiale de la lésion (Structural basis of UV DNA damage recognition by the DDB1-DDB2 complex 2009)
Ce basculement favorise le recrutement du complexe XPC-HR23B-CETN2.
b. Après cette étape, le complexe CRL4-DDB2, composé de CUL4A et RBX1 (activité ubiquitine-ligase) associés à DDB1 comme protéine adaptatrice et à DDB2 comme sous-unité de reconnaissance, prend le relais (Global-genome Nucleotide Excision Repair Controlled by Ubiquitin/Sumo Modifiers 2016).
2. Ce complexe catalyse une ubiquitination locale des histones H2, H3 et H4 ainsi que de la protéine XPC, et parfois de DDB2 elle-même, ce qui favorise l’ouverture locale de la chromatine et stabilise le recrutement du complexe XPC au site endommagé.
Réparation couplée à la transcription (TC-NER)
La réparation couplée à la transcription (TC-NER) intervient spécifiquement sur les gènes en cours de transcription.
Dans ce cas, la réparation est déclenchée lorsque l’ARN polymérase est bloquée par une lésion sur le brin matrice, ce qui recrute les facteurs du NER.
1. L'ARN polymérase bloquée est reconnue par le complexe CSA formé par les protéines CSA (ERCC8) et CSB (ERCC6), qui se fixent sur le complexe transcriptionnel arrêté et initient l’assemblage du complexe de réparation (Transcription-coupled DNA–protein crosslink repair by CSB and CRL4CSA-mediated degradation 2024).
- CSB possède une activité ATPase de type remodelage de chromatine qui contribue à stabiliser l’ARN polymérase II bloquée et peut favoriser son recul ou son déplacement, tout en facilitant le recrutement des facteurs du NER.
- Le complexe CSA fait partie d’un complexe ubiquitine-ligase de type CRL4-CSA, qui participe à la régulation des protéines associées à l’ARN polymérase arrêtée et facilite l’accès des facteurs de réparation au site de la lésion.
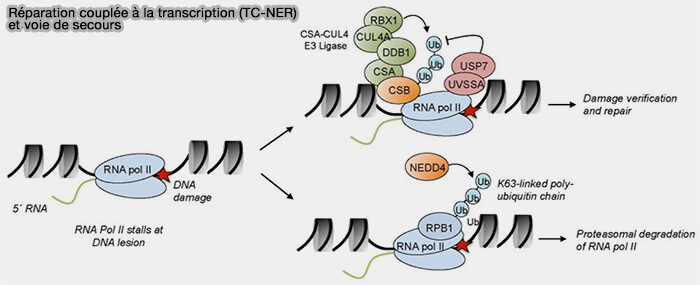
(Figure : vetopsy.fr d'après Stadler et Richly)
Remarque : lorsque la polymérase reste bloquée trop longtemps sur une lésion, la cellule élimine l’enzyme afin de libérer la région d’ADN et permettre la réparation (Regulation of DNA Repair Mechanisms: How the Chromatin Environment Regulates the DNA Damage Response 2017).
Cette dégradation constitue donc une voie de secours du TC-NER lorsque la polymérase ne peut pas être relancée.
2. Ces protéines permettent ensuite le recrutement des facteurs afin de former la bulle d’ADN simple brin nécessaire à l'excision.
Mécanisme commun d’excision et de réparation
Une fois la lésion reconnue, les deux sous-voies convergent vers un mécanisme commun d’excision et de réparation, le fragment éliminé contient généralement environ 24 à 32 nucléotides chez les eucaryotes.
1. Après la reconnaissance de la lésion, l’ADN est ouvert localement par le complexe TFIIH, composé principalement de 7 sous-unités (Core7), dont les hélicases XPB et XPD possèdent une activité hélicase dépendante de l’ATP (Dynamic conformational switching underlies TFIIH function in transcription and DNA repair and impacts genetic diseases 2023).
Ces protéines, qui remplissent des fonctions différentes, déroulent la double hélice autour du site endommagé en séparant les deux brins sur une région d’environ 20 à 30 nucléotides, ce qui élargit la bulle naissante d’ADN simple brin centrée sur la lésion.
(Figure : vetopsy.fr d'après Kim et coll)
a. XPB agit principalement comme moteur de translocation qui ouvre localement l’ADN ( mécanisme du domaine AAA).
mécanisme du domaine AAA).
b. XPD se déplace le long du brin endommagé et vérifie la présence effective des anomalies structurales détectées par XPC qui bloquent sa progression.
- Ces deux étapes de vérification évitent de couper l’ADN inutilement.
- Son canal de translocation est étroit et ne peut pas accommoder une base fortement modifiée ou une distorsion importante de l’ADN, de sorte que sa progression est physiquement bloquée lorsqu’elle rencontre la lésion, ce qui constitue le signal confirmant la présence effective du dommage.
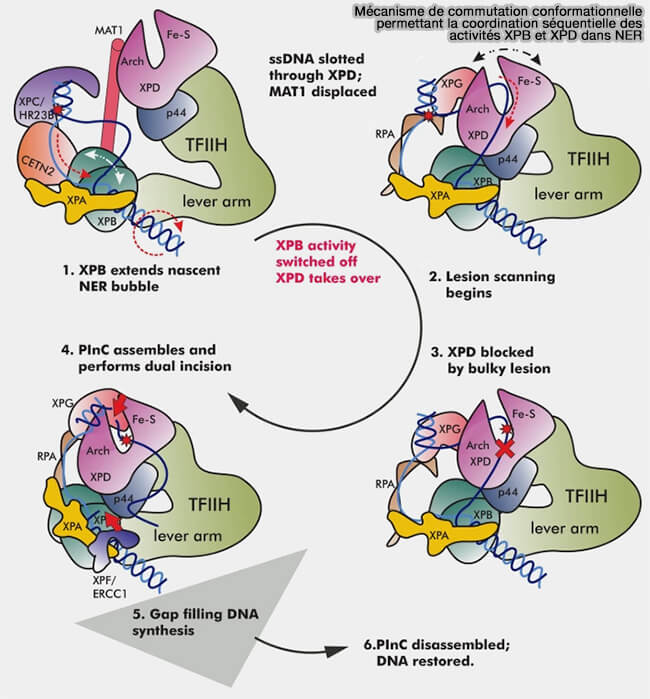
(Figure : vetopsy.fr d'après Yu et coll)
Remarque : le complexe TFIIH intervient également comme facteur général de transcription associé à l’ARN polymérase II, aux côtés de TFIIA, TFIIB, TFIID, TFIIE et TFIIF, dans lequel il participe notamment à l’ouverture locale de l’ADN au niveau du promoteur et à l’initiation de la transcription.
2. La région ouverte est ensuite stabilisée par les protéines XPA et RPA.
a. XPA, en se fixant à la jonction entre l’ADN simple brin et l’ADN double brin dans la bulle de réparation, stabilise la structure ouverte de l’ADN.
Elle interagit directement avec plusieurs composants du NER, notamment RPA, TFIIH et le complexe XPF-ERCC1, ce qui permet d’organiser l’assemblage du complexe d’incision et de positionner correctement les endonucléases de part et d’autre de la lésion.
b. RPA se fixe sur l’ADN simple brin afin de stabiliser la bulle et d’empêcher la réhybridation des brins pendant les étapes d’incision (stabilisation de l'ADN simple brin par RPA)
3. Ces étapes sont résumées dans la figure ci-dessous dans laquelle (Lesion recognition by XPC, TFIIH and XPA in DNA Excision Repair 2023) :
- C7CD montre le recrutement de TFIH : XPC est lié à l’ADN endommagé, TFIIH est recruté (Core7), la lésion est encore positionnée près de XPC qui maintient l’ADN ouvert, la lésion n'est pas encore vérifiée par XPD.
- C7CAD montre le repositionnement du complexe : l'insertion de XPA dans le complexe stabilise la bulle d’ADN, repositionne TFIIH sur l’ADN et rapproche l’ADN de l’hélicase XPD.
- C7AD montre la préparation de la vérification par XPD : XPC s'est détaché lorsque XPA, s’est correctement fixé et le complexe actif est alors TFIIH + XPA + AD, XPD qui se déplace de le long du brin endommagé (5' → 3') pour agir comme capteur de lésion.
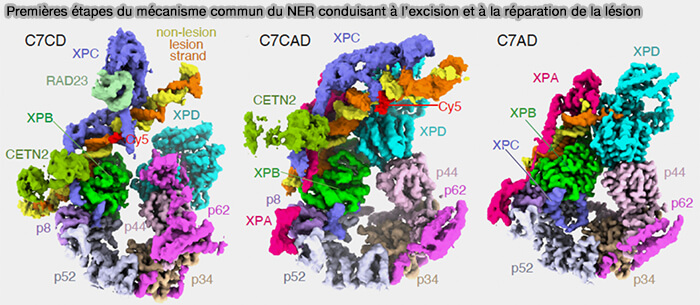
(Figure : vetopsy.fr d'après Kim et coll)
3. Deux endonucléases réalisent ensuite des incisions précises de part et d’autre de la lésion, ce qui permet l’excision d’un oligonucléotide contenant la lésion généralement long d’environ 24 à 32 nucléotides chez les eucaryotes (Nucleotide Excision Repair: From Molecular Defects to Neurological Abnormalities 2021).
a. Le complexe XPF-ERCC1 coupe le brin endommagé du côté 5' de la lésion.
- XPF n’est actif qu’en association avec la protéine ERCC1, avec laquelle il forme un hétérodimère stable.
- ERCC1 participe à la reconnaissance de la structure ADN et positionne correctement l’activité nucléase de XPF.
b. L’endonucléase XPG réalise l’incision du côté 3' du site endommagé.
Contrairement à XPF, XPG possède à elle seule le domaine catalytique nucléase et peut agir comme enzyme monomérique, bien qu’elle interagisse avec plusieurs protéines du NER, notamment TFIIH et RPA, pour être correctement positionnée sur l’ADN.
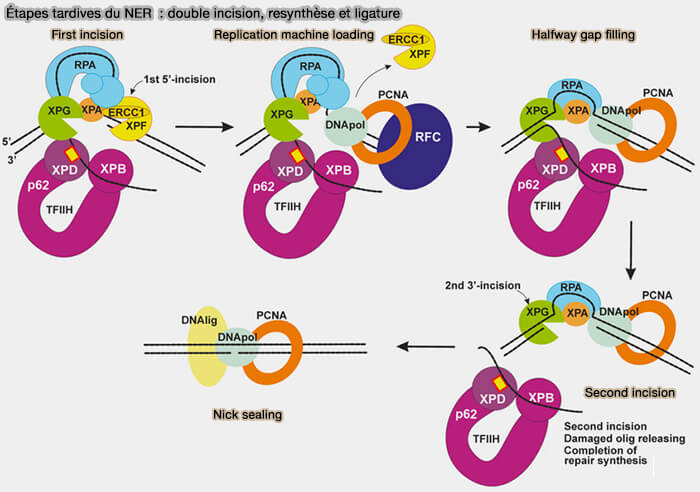
(Figure : vetopsy.fr d'après Krasilova et coll)
4. Après l’excision de l’oligonucléotide contenant la lésion, l’espace laissé dans le brin d’ADN est comblé par une synthèse de réparation en utilisant le brin complémentaire intact comme matrice, réalisée par des ADN polymérases, principalement les polymérases δ ou ε associées au facteur de réplication PCNA récruté par RFC.
5. Enfin, une ADN ligase rétablit la continuité du squelette phosphodiester en reliant les extrémités du brin néosynthétisé au reste de la molécule d’ADN, ce qui restaure l’intégrité structurale de la double hélice.
- La ligase I intervient principalement lors de la réparation associée aux facteurs de réplication, PCNA et RFC ( maturation des fragments d'Okazaki).
- Le complexe ligase III-XRCC1 agit surtout dans des contextes de réparation indépendants de la réplication, XRCC1 servant de protéine d’échafaudage qui stabilise la ligase III et coordonne les interactions avec d’autres protéines de réparation.
Autres réparations de l'ADN
BiochimieChimie organiqueBioénergétiqueAcides nucléiquesNucléotidesADNStructureSynthèseOrganisation du génomeRéplicationTranscriptionÉpigénétiqueRéparation de l'ADNARNChromatineNucléosomesHistonesChromosomesProtidesGlucidesLipidesEnzymesCoenzymesVitaminesHormonesComposés inorganiquesTransport membranaireMoteurs moléculairesVoies de signalisation