Les structures cristallines du complexe Syt1-SNARE révèlent trois interfaces entre le complexe SNARE et les domaines C2A et C2B de Syt1, i.e. appelées primaire, secondaire et tertiaire.
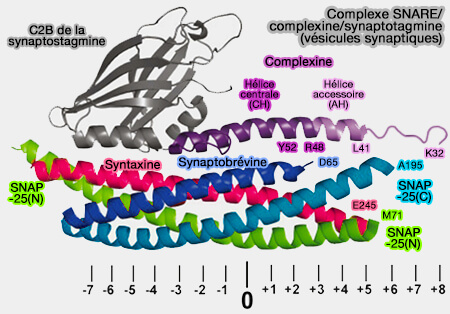
Structure cristalline du complexe Syt1-SNARE
(Figure : vetopsy.fr d'après Zhou et coll)
Interface primaire
1. L'interface primaire, la plus grande, formée entre le domaine C2B de Syt1 et le complexe SNARE, en particulier avec SNAP-25 et la syntaxine 1 (Stx1), est divisée en deux régions dominées par les interactions polaires.
La région I montre l'interaction entre les résidus de SNAP-25, i.e. N159, H162, M163 et D166, et ceux de Syt1, i.e. E295, K297, N336 et Y338.
La région II comprend l'interaction entre les résidus de Stx1, i.e. D231, E234, E238 et ceux de Syt1, i.e. R281, K288, R398 et R399, i.e. ces deux derniers résidus correspondent à l'apex d'arginine impliqué dans la fusion vésiculaire.
Les deux régions polaires sont reliées par un patch hydrophobe, i.e. I44, L47 et V48 de SNAP-25 et V292, L294, A402 de Syt1.
Le complexe SNARE est chargé positivement et Syt1 est chargé négativement dans la région I, alors que c'est l'inverse pour la région II.
2. Cinq résidus clés forment le site de liaison primaire, basé sur des données structurelles, i.e. cette interface est hautement conservée et essentielle pour la fonction Syt1.
b. La perturbation de la liaison SNARE sur ce site mime les mutants nuls de Syt1 parfaitement, de manière critique, y compris la perte de fusion synchrone et des taux élevés de libération asynchrone et spontanée.
Ainsi, Syt1 et Cplx se lient la surface externe du complexe SNARE pour former une hélice α scindée mais continue, i.e. Syt1 et Cplx pourraient former une seule unité de régulation qui réduit la barrière d'énergie nécessaire pour la fermeture complète du complexe SNARE.
Résumé des interfaces
Deux molécules Syt1 interagissent simultanément avec deux interfaces de liaison sur les côtés opposés du complexe SNARE :
Les deux interfaces de liaison sont essentielles pour la libération de neurotransmetteurs déclenchée par Ca++.
Lorsque l'interface tripartite est perturbée alors que l'interface primaire est intacte, ou vice versa, on assiste à une réduction de la libération évoquée.
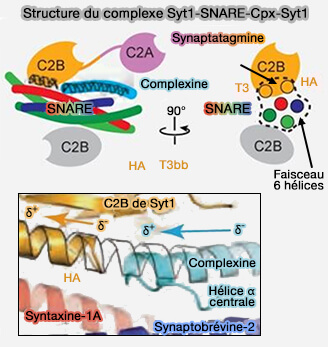
Complexe Syt1-SNARE-Cplx-Syt1
(Figure : vetopsy.fr d'après Zhou et coll)
1. L'interface primaire Syt1-SNARE est spécifique aux capteurs Ca++ rapides (Syt1, Syt2 et Syt9).
2. L'interface tripartite pourrait être une interface plus générale impliquant d'autres synaptotagmines compte tenu de la conservation de la séquence des hélices T3 et HA parmi tous les domaines C2B des synaptotagmines, excepté les domaines C2A des synaptotagmines ou C2 de Munc13.
La structure du complexe Syt1-SNARE-Cplx-Syt1 serait l'état de pré-fusion du complexe.
L'interface tripartite active et verrouille en même temps le complexe, gardant le complexe trans-SNARE sous tension à moitié zippé et les membranes séparées, empêchant ainsi la fusion des membranes.
Conformément à ce modèle de l'état amorcé, l'insertion constitutive de Cplx dans le complexe SNARE inhibe la libération des vésicules synaptiques, et l'inclusion de Cplx augmente la séparation entre les membranes (Morphologies of synaptic protein membrane fusion interfaces 2017).










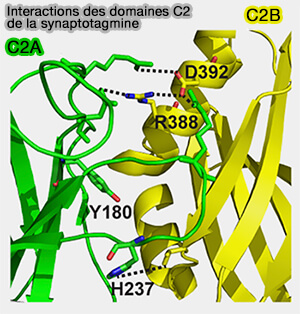
 mécanisme de l'otoferline).
mécanisme de l'otoferline).