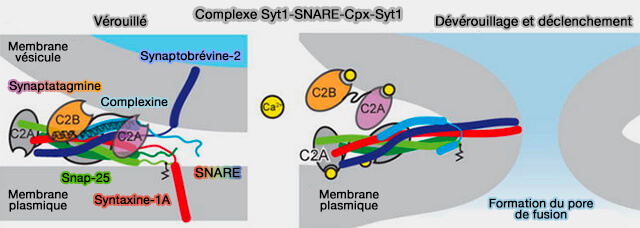
Différents modèles controversés, mais qui peuvent se chevaucher partiellement, ont été formulés pour expliquer comment la synaptotagmine-1 (Syt1) et la complexine (Cplx) couplent la liaison Ca++ à la fusion membranaire ( modèles de régulation des protéines SNARE).
Le modèle décrit ci-dessous correspondrait à un mélange des modèles suivants :
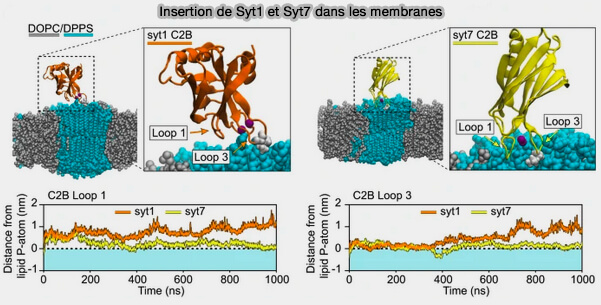
a. Les boucles 1 ou 3 établissent plus de contacts avec la membrane que la boucle 2, excepté dans C2A ou C2B de Syt7 chez lesquels elle entre en contact par trois résidus basiques consécutifs avec les phospholipides membranaires.
Liaison membranaire de C2A de Syt1 et de Syt7
(Figure : vetopsy.fr d'après Vermaas et Tajkhorshid)
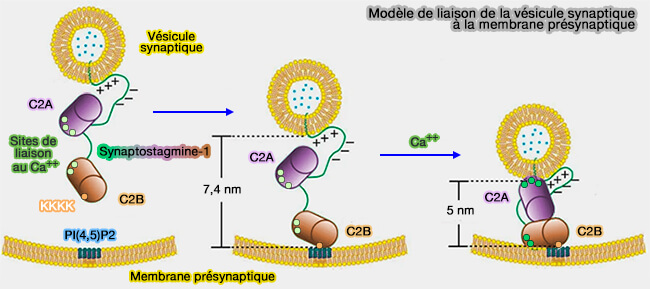
b. Syt1 lie Ca++ et relie les deux bicouches de sorte que C2A et C2B se lient aux surfaces opposées de la bicouche via leurs boucles de liaison Ca++.
Syt1 pourrait modifier la distance membrane vésiculaire/membrane plasmique, et il est concevable que cet événement puisse déclencher l'assemblage SNARE ( dévérrouillage du complexe SNARE).
Modèle de liaison de la vésicule synaptique à la membrane présynaptique
(Figure : vetopsy.fr d'après Lin et coll)
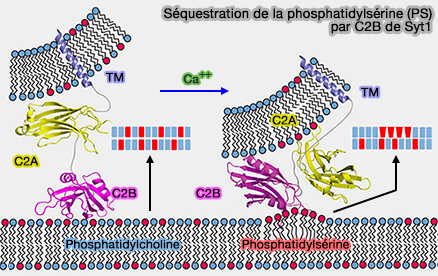
moduler l'association membranaire d'autres protéines, telles que Munc13 ou les régions juxta-membranaires des SNARE,
conduire à une déformation de courbure positive.
Séquestration de la phosphatidylsérine (PS) par C2B de Syt1
(Figure : vetopsy.fr d'après Lai et coll)
2. Simultanément, le patch polybasique améliore son affinité pour PI(4,5)P2 ou PIP2, conduisant à une pénétration plus profonde du site de liaison Ca++ dans la bicouche.
Les fonctions d'amarrage/amorçage du patch de lysine polybasique facilitent la libération, mais ne sont pas essentielles puisque les sites de liaison au Ca++ restent intacts et permettent toujours la liaison à la PS et l'insertion de la membrane.
3. La liaison de Ca++ augmente le temps de séjour de Syt1 sur la membrane résultant d'une stabilité cinétique élevée du complexe Syt-1-membrane, en permettant un contact plus intime avec la bicouche plutôt qu'en favorisant une amélioration de l'association synaptotagmine/membrane.
Lors de la liaison du calcium, les deux protéines pénètrent partiellement les bicouches lipidiques qui portent des phospholipides anioniques.
1. Les interactions de Syt1 avec la membrane sont grandement influencées par l'ordre membranaire.
L'emballage serré de la phosphatidylsérine inhibe la liaison en raison d'une altération de la pénétration de la membrane.
Insertion de Syt1 et Syt7 dans les membranes
(Figure : vetopsy.fr d'après Courtney et coll)
2. En revanche, Syt7 a une activité de liaison et de pénétration de la membrane plus robuste quelle que soit la structure de la chaîne acyle des phospholipides.
Elle permet ainsi la formation de pores plus grands et plus stables que ceux de Syt1.
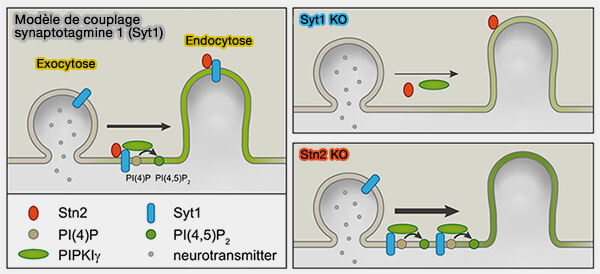
Elle recrute PIPKIγ, i.e. enzyme de synthèse de PI(4,5)P2, sur les sites endocytaires.
PI(4,5)P2 est synthétisé localement dans la zone péri-active.
2. La synthèse locale de PI(4,5)P2 déclenchée par Syt1 est probablement en synergie avec d'autres mécanismes potentiels de couplage exo-endocytaire (couplage exocytose/endocytose).
Les queues lipidiques du feuillet proximal interagissent avec l'hélice déplacée et deviennent désordonnées.
Hélice HB de C2B de Syt1
(Figure : vetopsy.fr d'après Wu et Schulten)
Les queues du feuillet distal, pour rester en contact avec le feuillet proximal, s'étirent et s'ordonnent.
2. Les deux résidus hydrophobes, localisés à l'extrémité des boucles de liaison Ca++ de chaque domaine C2 de Syt1, i.e M173, F234, V304 et I367, sont partiellement insérés dans le feuillet interne de la membrane plasmique (10 Å de profondeur) et peuvent induire une déformation locale de la membrane plasmique.
Ce processus entraîne un déséquilibre de pression à travers la membrane et provoque ainsi une flexion de la membrane.
La barrière énergétique à la formation de la tige diminue également, car la courbure positive pourrait réduire l'énergie de flexion de la membrane nécessaire pour former une tige.
La liaison membranaire des domaines C2 est nécessaire pour initier la fusion, mais les interactions membranaires créées par l'apex d'arginine sont nécessaires pour en déterminer le bon moment et l'ouverture rapide du pore de fusion (ouverture du pore de fusion).
Les changements conformationnels de Syt1 dépendants de Ca++ délogeraient Cplx pour faciliter la conversion du complexe trans-SNARE en complexe cis-SNARE pour conduire la réaction de fusion finale.
Le déplacement de Cplx fournirait des sites de liaison pour α-SNAP afin d'initier le désassemblage ultérieur du complexe SNARE médié par la NSF.
Rôle de C2A de Syt1 dans la liaison membranaire
(Figure : vetopsy.fr d'après Bowers et coll)
Ce changement morphologique de la membrane plasmique rapproche les membranes, plus près que la distance critique (0,9 nm),pour favoriser la formation des tiges de fusion qui conduit ensuite à l'ouverture des pores de fusion.
Remarque : plusieurs complexes synaptiques impliqués pourraient potentiellement interagir les uns avec les autres, i.e. le domaine C2B de Syt1 pourrait relier deux complexes SNARE via les interfaces primaires et tripartites.




 modèles de régulation des protéines SNARE).
modèles de régulation des protéines SNARE).