


Fission membranaire
Mécanismes
- Biologie cellulaire et moléculaire
- Constituants de la cellule
- Transport membranaire
- Moteurs moléculaires
- Voies de signalisation

La fission membranaire passe par l'étape de formation du col vésiculaire, par l'étape intermédiaire d'hémifission et par la formation de deux membranes distinctes.
Machinerie de fission
La fission membranaire passe par les étapes suivantes (Protein Amphipathic Helix Insertion: A Mechanism to Induce Membrane Fission 2019) :
- l'étape intermédiaire du col de la membrane,
- l'étape intermédiaire d'hémifission,
- la formation de deux membranes distinctes.

Les étapes du cou et de l'hémifission du processus de fission membranaire ressemblent aux intermédiaires des pores et du diaphragme d'hémifusion du processus de fusion membranaire ( mécanisme de fusion membranaire).
mécanisme de fusion membranaire).
Vous pouvez lire :
- pour le détail du rôle de la dynamine : The role of scaffold reshaping and disassembly in dynamin driven membrane fission (2018),
- pour les discussions détaillées des modèles et sur les controverses : Membrane fission by dynamin: what we know and what we need to know (2016).

Il ne faut pas oublier que les mécanismes de la fusion sont encore étudiés à l'heure actuelle, car des zones de flou subsistent !
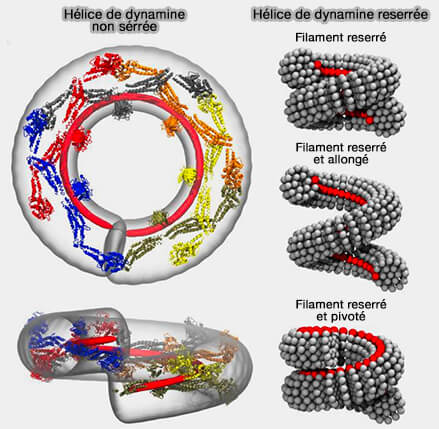
(Figure : vetopsy.fr d'après Pannuzzo et coll)
Fission et dynamine
Nous prendrons comme exemple le rôle de la dynamine dans la fission.
Constriction du
cou de la vésicule
1. La vésicule subit une constriction au niveau du col membranaire suivant différents processus.
Pour la dynamine, il existe deux modèles qui ont chacun des inconvénients :
Ces deux modèles sont étudiés dans un chapitre spécial.
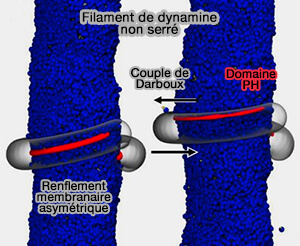
(Figure : vetopsy.fr d'après Pannuzzo et coll)
2. L'hélice de dynamine peut effectivement être décrite comme une rotation nette de l'échafaudage hélicoïdal autour de son axe longitudinal local, même si le noyau du filament protéique ne tourne pas réellement.
- Les domaines PH, qui attachent l'hélice de dynamine à la membrane, doivent s'incliner pour faire pivoter la bande adhésive, i.e. les domaines PH, loin du substrat.
- Un filament non resserré reposant sur une membrane de rayon correspondant crée un couple de Darboux tangentiel sur la membrane, i.e. cadre de Darboux, une fois que la bande d'adhérence, i.e. les domaines PH, tourne, induisant un renflement asymétrique de la membrane.
2. Une simulation informatique qui reproduit fidèlement la géométrie et l'élasticité de la membrane et de la dynamine a été utilisée pour tester différentes façons dont la protéine pourrait fonctionner (The role of scaffold reshaping and disassembly in dynamin driven membrane fission 2018).
- Le premier test comportait une simple constriction, où la spirale de dynamine se contracte autour de la membrane pour la pincer, i.e. la vésicule ne s'est séparée de la membrane qu'après une constriction invraisemblablement serrée.
- Le deuxième test ajoute au premier un allongement de la spirale ainsi que la réduction de son diamètre, i.e. ce qui n'a fait que réduire la capacité de la protéine à se détacher de la membrane.
- Le test final combinait constriction et rotation, la dynamine " tournoyant " en appuyant sur le col de la vésicule, i.e. le détachement de la vésicule a été efficace une fois la spirale de dynamine démontée.

3. La constriction et le désassemblage de l'échafaudage de dynamine seraient concomitants, i.e. les filaments courts induisent l'hémifission plus efficacement que les longs (Dynamin's helical geometry does not destabilize membranes during fission 2018).
- Cette vidéo est identique à la précédente, sauf que le filament se dépolymérise simultanément pendant la constriction et la rotation.
- En conséquence, une fois l'hémifission installée, les deux défauts ponctuels aux extrémités de la micelle d'hémifission cylindrique résultante ne sont plus éloignés par l'échafaudage maintenant beaucoup plus court, permettant aux défauts de s'annihiler et donc de terminer la fission.
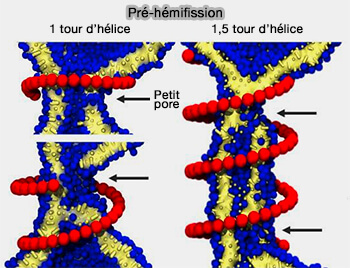
(Figure : vetopsy.fr d'après Pannuzzo et coll)
Lors de l'hydrolyse du GTP, l'échafaudage semble s'ajuster à une longueur optimale pour la fission, i.e. environ un tour complet (Dynamin-driven membrane fission reveals catalytic principle of membrane remodeling 2013).
Hémifission
1. L'hémifission est un état intermédiaire où la monocouche proximale (également appelée monocouche de contact) de la bicouche formant le col fusionne, séparant ainsi le volume interne en deux parties, tandis que la monocouche distale reste continue (A hemi-fission intermediate links two mechanistically distinct stages of membrane fission 2015).
Cependant, deux résultats d'expériences in vitro ont été en désaccord apparent avec cette vue la plus simple.
- Premièrement, la constriction de la dynamine est nécessaire, mais pas suffisante pour la fission (A Dynamin Mutant Defines a Superconstricted Prefission State 2014). La dynamine pourrait resserrer le tubule membranaire à une taille qui atteint spontanément l'hémi-fission d'une manière dépendante de la tension et de la rigidité qui dépendent de la physique membranaire.
- Deuxièmement, l'hydrolyse du GTP déclenche une dépolymérisation partielle de la couche de dynamine.
L'induction de l'hémifission reste encore floue expérimentalement.
2. La constriction seule, jusqu'au plus petit rayon luminal observé expérimentalement, ne suffit pas quel que soit le modèle, i.e. si l'état resserré est atteint :
- passivement (modèle en deux étapes),
- activement (modèle constrictase).
a. L'inclinaison du domaine PH apparaîtrait comme un moyen de catalyser l'hémifission, i.e. l'énergie de l'hydrolyse du GTP pourrait au moins en partie être utilisée pour entraîner ce changement conformationnel (A Dynamin Mutant Defines a Superconstricted Prefission State 2014).
Le remplacement du domaine PH par un simple motif de liaison ralentit fortement le taux de fission (The pleckstrin-homology domain of dynamin is dispensable for membrane constriction and fission 2017).
b. Ce domaine contient une boucle amphipathique assez courte qui pourrait s'enfoncer dans la membrane pour la resserrer davantage (Membrane Insertion of the Pleckstrin Homology Domain Variable Loop 1 Is Critical for Dynamin-catalyzed Vesicle Scission 2009).
- Cependant, la position de cette boucle, éloignée de la poche de liaison PI(4,5)P2 ou PIP2 dans la structure PH, ne permet pas une insertion dans la membrane sans libérer son lien avec PIP2.
- De plus, la boucle (quelques acides aminés) est si courte que l'on peut s'interroger sur le fait qu'elle puisse générer suffisamment de courbure pour resserrer davantage la membrane.
c. Toutefois, les forces générées par l'interaction du dimère GTPase sont transférées à travers la tige vers le domaine PH et de là vers la membrane (Cryo-EM of the dynamin polymer assembled on lipid membrane 2018).
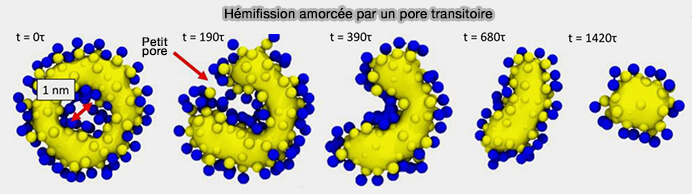
(Figure : vetopsy.fr d'après Pannuzzo et coll)
3. Il semblerait que l'hémifission commence par la formation d'un petit pore transitoire perforant le cou resserré.
- Or, la fission est censée être sans fuite et l'existence d'un intermédiaire d'hémifission est généralement considérée comme excluant la nécessité ou même la possibilité de pores.
- Les pores de fuite observés sont partiellement couverts par l'échafaudage de dynamine et, dans des conditions physiologiques, par des protéines supplémentaires à domaines BAR par exemple, ce qui pourrait diminuer significativement la conductance effective d'un tel pore.
Cependant, le feuillet interne d'un tube doit changer de topologie, posant la question de savoir comment un filament uniquement en contact avec le feuillet externe peut favoriser cette transition.
- Le filament amorcerait la formation d'un petit pore qui s'élargit autour de la circonférence du cou, tandis que ses bords supérieur et inférieur tirent radialement vers l'intérieur et fusionnent avec la membrane intacte.
- Cela crée deux lignes de défaut au niveau desquelles trois bicouches se rencontrent, et leur contraction entraîne la singularité des points supérieur et inférieur de la micelle cylindrique de l'état d'hémifission.
Cette séquence d'événements n'implique jamais un grand pore, à condition que le filament reste suffisamment court, ce qui suggère qu'une coordination étroite entre la constriction, la rotation et éventuellement le désassemblage concomitant rend la fission plus efficace.
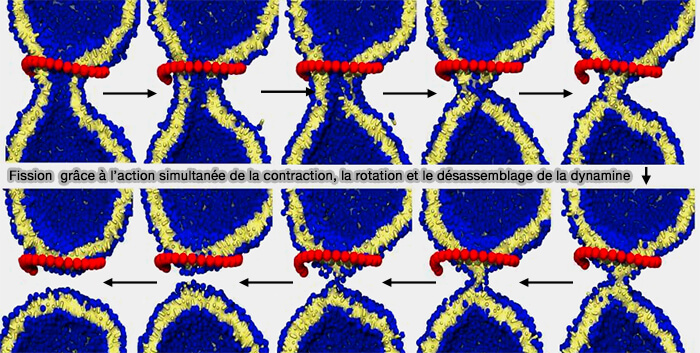
(Figure : vetopsy.fr d'après Pannuzzo et coll)
Autres protéines de fission
Le modèle décrit ci-dessus pourrait bien être aussi valable pour d'autres protéines de fission (The role of scaffold reshaping and disassembly in dynamin driven membrane fission 2018).
1. Ce peut être sûrement le cas des autres membres de la superfamille de la dynamine (DSP), qui partagent de nombreuses propriétés avec la dynamine, en particulier vraisemblablement l'oligomérisation en hélice.
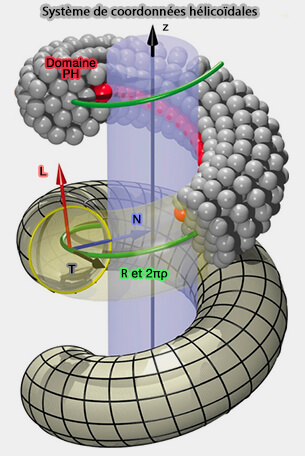
(Figure : vetopsy.fr d'après Pannuzzo et coll)
Les couples de Darboux sur la membrane peuvent provoquer la transition vers l'état d'hémifission, et donc ils fournissent un moyen de déclencher une transition topologique qui est différente des notions de constriction ou d'allongement.
2. La fission inversée par le complexe ESCRT-III implique l'adsorption et la polymérisation d'un filament hélicoïdal à l'intérieur du col à sectionner, dont le mode de fonctionnement est beaucoup moins bien compris que celui de la dynamine classique.
a. Les modèles actuels reposent sur l'adhérence et/ou les changements géométriques des filaments en spirale, qui entraînent soit des forces directes, soit des contraintes induites par les limites au niveau du site de contact (modèles de remodelage membranaire et de scission par ESCRT).
b. Mais les réarrangements géométriques d'un filament élastique incurvé avec une rigidité de torsion finie, dont le cadre matériel est épinglé à la membrane (Environmental bias and elastic curves on surfaces 2014), entraînent presque invariablement :
- des couples de Darboux supplémentaires, ainsi que
- des analogues induits par la torsion (Shape Selection of Surface-Bound Helical Filaments: Biopolymers on Curved Membranes 2016).
Fission des différents compartiments membranaires
La fission intervient dans :
- l'endocytose,
- les endosomes,
- les lysosomes,
- les mitochondries…
Biologie cellulaire et moléculaireConstituants de la celluleTransport membranaireTransports sans mouvements membranairesTransports passifsTransports actifsTransports avec mouvements membranaires : trafic vésiculaireFusion/fission membranaireEndocytoseVoie sécrétoireCanaux ioniquesTransporteursUniportsPompesCo-transporteursMoteurs moléculairesVoies de signalisation