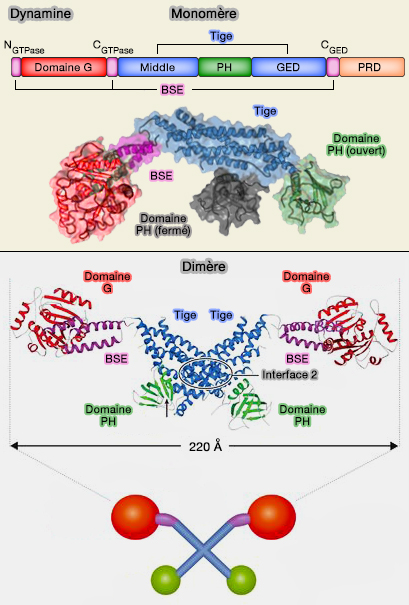
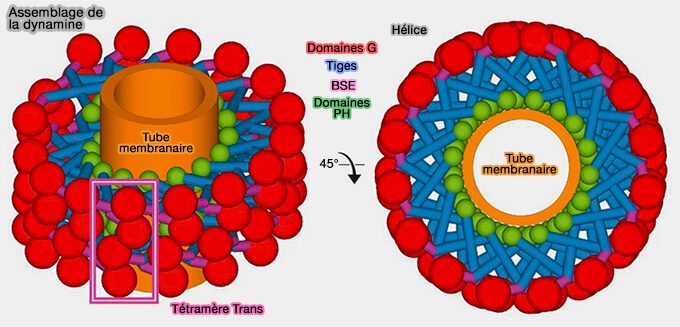
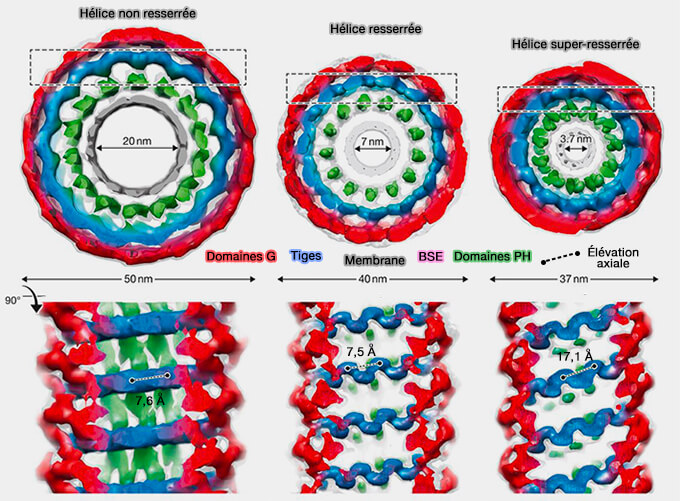
Hélice de dynamine
(Figure : vetopsy.fr d'après Antonny et coll)
En l'absence de nucléotide, la dynamine s'assemble en une couche hélicoïdale de 50 nm de diamètre extérieur avec un pas hélicoïdal entre 10 et 20 nm entourant un tube de membrane de 10 nm de rayon.
Hélice de dynamine
Figure : vetopsy.fr d'après Antonny et coll)
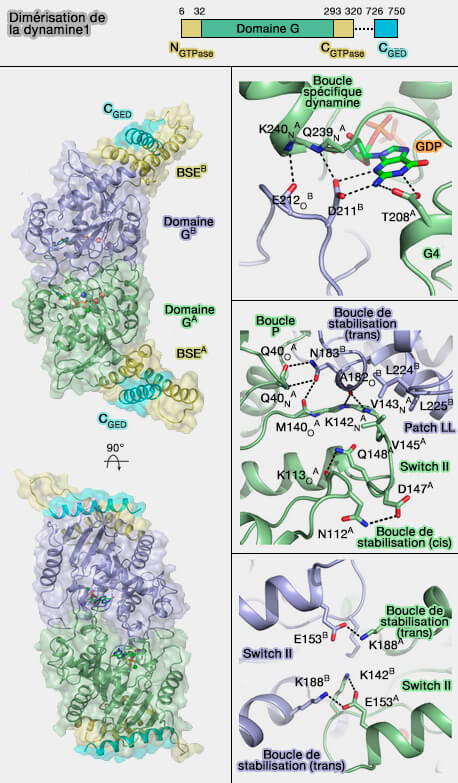
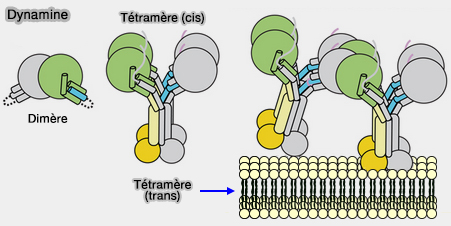
Dimérisation
L'oligomérisation de " base " est la dimérisation.
Dimérisation de la dynamine
(Figure : vetopsy.fr d'après Chappie et coll)
Les polymères de dynamine sont des dimères anti-parallèles :
2. Cette dimérisation, grâce à des liaisons hydrogènes et des ponts salins, fait intervenir :
la boucle de stabilisation cis (205-216) et le motif TKLD (205-208) du domaine G4 ;
la boucle spécifique de la dynamine (DSL, dynamin specific loop, 236-249) ;
la boucle de stabilsation trans (176-188) avec le switch II (136-153), renforcé par les interactions entre V145 du switch II et les leucines 224 et 225 (patch LL) et par une liaison hydrogène entre N183 et les atomes de la chaîne principale de Q40 dans la boucle P.
Oligomérisation d'ordre supérieur
1. Les interactions entre les dimères de la tige, par les interface 1 et 3, assurent l'auto-assemblage (appelé souvent oligomérisation d'ordre supérieur) hélicoïdal qui produisent le polymère de la taille attendue.
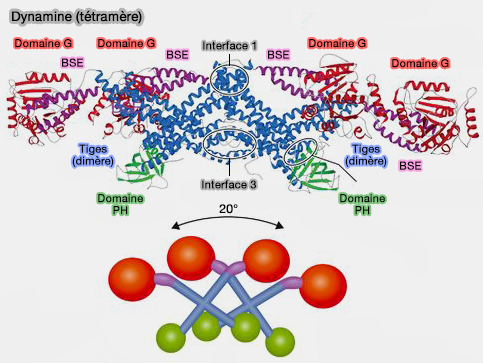
Tétramère de dynamine
(Figure : vetopsy.fr d'après Antonny et coll)
2. Dans les dynamines non assemblées, le domaine PH est replié sur la tige, induisant une conformation " fermée " auto-inhibée qui empêche l'oligomérisation au-delà de l'état tétramérique.
2. L'activité basale de la GTPase basale (~ 0,5-2/minute min-1) émane des tétramères qui ne peuvent s'auto-assembler en hélice par des interactions auto-inhibitrices entre les domaines PH et la tige (Crystal structure of the dynamin tetramer 2015).
Dimérisation et tétramérisation de la dynamine
(Figure : vetopsy.fr d'après Chappie et coll)
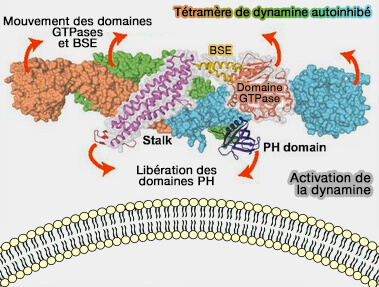
Lorsque le GTP est lié (l'état prédominant dans la cellule), le BSE adopte une conformation ouverte et se projette vers l'extérieur, les domaines G étant appariés à l'intérieur.
Lors de l'hydrolyse du GTP, le BSE adopte une conformation fermée en se repliant contre la face postérieure des domaines G.
Ce changement conformationnel provoque des réarrangements spécifiques dans la tige et/ou dans les domaines en interactions avec la membrane, i.e. facilite sa constriction et la fission.
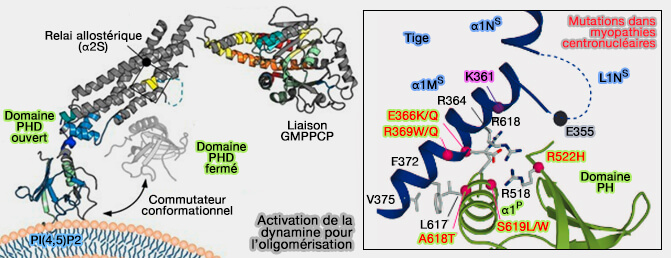
Le domaine PH de la dynamine, qui s'insère partiellement dans la bicouche membranaire via des interactions hydrophobes, facilite probablement la déstabilisation membranaire lors de la fission.
En effet, Les DSP qui ne possèdent pas ce domaine, contractent, mais ne rompentt pas les membranes.
Auto-assemblage
L'auto-assemblage d'ordre supérieur en polymères hélicoïdaux autour des matrices membranaires contenant du phosphatidylinositol-4,5-bisphosphate stimule l'hydrolyse du GTP d'au moins 200 fois.
1. L'oligomère hélicoïdal de dynamine a la capacité à se contracter en présence de GTP.
Cet état super-resserré, i.e. dans des concentrations super-saturantes de GTP et en présence de la dynamine mutante à affinité réduite pour le GTP inhibant partiellement sa GTPase et son activité de fission, est obtenu par l'assemblage d'une hélice à deux départs.
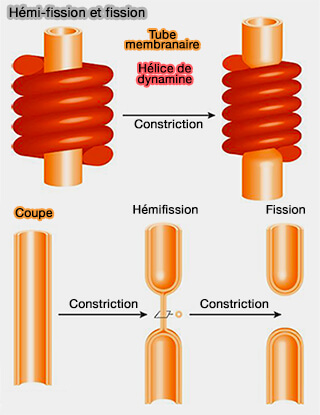
Dans la première description de ce modèle, la dynamine, pendant l'hydrolyse du GTP, se contracterait jusqu'à ce que le col de la membrane atteigne l'état d'hémi-fission et soit complètement rompu.
L'hémi-fission apparaît quand le feuillet interne du tube disparaît, laissant un col de liaison constitué d'une seule monocouche lipidique enveloppée dans une micelle cylindrique.
Cependant, deux résultats d'expériences in vitro ont été en désaccord apparent avec cette vue la plus simple.
Premièrement, la constriction de la dynamine est nécessaire, mais pas suffisante pour la fission (A Dynamin Mutant Defines a Superconstricted Prefission State 2014). La dynamine pourrait resserrer le tubule membranaire à une taille qui atteint spontanément l'hémi-fission d'une manière dépendante de la tension et de la rigidité qui dépendent de la physique membranaire.
Deuxièmement, l'hydrolyse du GTP déclenche une dépolymérisation partielle de la couche de dynamine.
Ainsi, la constriction par la dynamine peut ne pas être suffisante à elle seule pour provoquer la fission de la membrane, mais plutôt la dynamine pourrait resserrer le tubule membranaire à une taille qui atteint spontanément l'hémi-fission d'une manière dépendante de la tension et de la rigidité.