Les ATPases de type P (P-ATPases) forment une famille de protéines membranaires qui utilisent l'énergie de l'hydrolyse de l'ATP pour transporter les ions et les lipides à travers les membranes biologiques.
Ces pompes, parfois E1E2-ATPases se trouvent chez les procaryotes, les archées et les eucaryotes.
Les P-ATPases se distinguent des autres transporteurs dépendants de l'ATP, i.e. les ATPases à moteur rotatifs, F-ATPases, V-ATPases, et les transporteurs ABC, par la présence d'un aspartate conservé qui subit une phosphorylation transitoire au cours du cycle ATP, d'où le nome de type P.
Les membres de cette famille génèrent et maintiennent des gradients électrochimiques cruciaux à travers les membranes cellulaires, en transloquant les cations, les métaux lourds et les lipides.
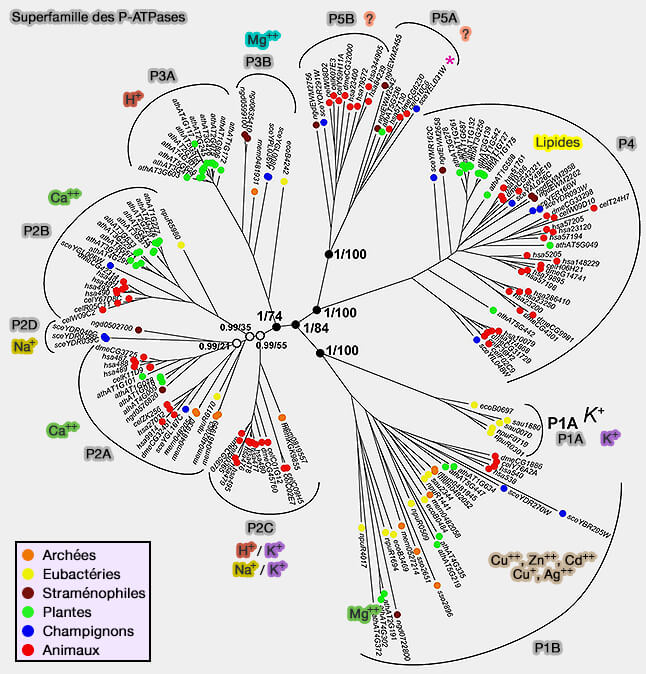
Classification des P-ATPases
Les ATPases de type P, qui comprennent plus de 20 familles différentes, forment cinq classes (Evolution of P-type ATPases 1998).
Les P2C-ATPases transportent des ions monovalents et comprennent les Na+/K+-ATPases et les H+/K+-ATPases.
La pompe sodium-potassium (Na+/K+-ATPase) des animaux et la H+/K+-ATPases des plantes et des champignons maintiennent le potentiel de la membrane plasmique dans toutes les cellules eucaryotes, qui est basé sur les différentes concentrations d'ions des deux côtés de la membrane.
Les P2D-ATPases se trouvent dans les champignons et transportent Na+.
3. Les P3-ATPases
Les P3A-ATPases transportent H+.
Les P3B-ATPases transportent Mg++.
4. Les P4-ATPases sont uniques en ce sens qu'elles sont les seules à transporter des lipides, i.e. des phospholipides, substrat dix fois plus gros que les ions, à travers les membranes.
Les P4-ATPases sont étudiées dans un chapitre spécifique.
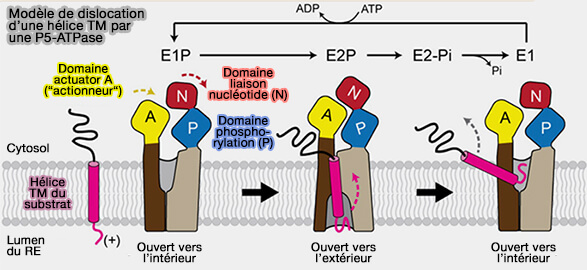
Modèle de dislocation d’une hélice TM par une P5-ATPase
(Figure : vetopsy.fr d'après Mckenna et coll)
b. Le mécanisme serait le suivant.
Pendant la transition E1P-à-E2P, le domaine N tourne et le domaine A se rapproche de l'interface N-P, provoquant la conversion de la poche de liaison au substrat ouverte vers l'intérieur du domaine transmembranaire à la conformation ouverte vers l'extérieur.
Dans ce modèle, une hélice TM de substrat avec un segment luminal court et préférentiellement chargé positivement se lierait à la poche ouverte vers l'extérieur et la transition E2P à E1 ferait basculer la TM par un interrupteur de l'ouverture vers l'extérieur vers l'intérieur.
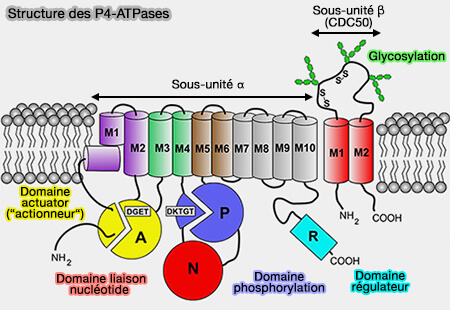
Structure des P4-ATPases
(Figure : vetopsy.fr d'après Andersen et coll)
Elles peuvent également contribuer à la spécificité du substrat en variant d'une protéine à l'autre.
b. Deux particularités dans les hélices TM contribuent à la formation d'une poche de liaison intégrée à la membrane pour les cations du substrat.
2. On peut séparer le domaine M en :
sous-domaine N-terminal, i.e. NTM (formé par M1-M4a),
sous-domaines C-terminal, i.e. CTM (M4b-M10).
a. En effet, le site de liaison au substrat est situé entre NTM et CTM, bien qu'il soit davantage associé à CTM.
b. Les changements conformationnels à l'interface NTM/CTM au cours du cycle E1-E2 permettent un accès alterné de ce site de liaison au cytosol ou à l'espace extracellulaire/luminal.
Domaines cytoplasmiques
Les domaines cytoplasmiques comprennent trois domaines.
1. un domaine actuator (A), actionneur en français, i.e. qui transforme l’énergie qui lui est fournie en un phénomène physique.
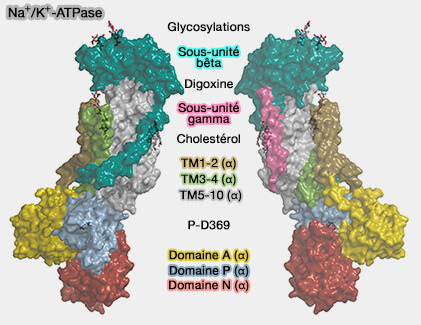
Structure de Na+/K+-ATPase
(Figure : vetopsy.fr d'après Clausen et coll)
Le domaine A cytosolique représente une insertion dans le NTM, connecté aux hélices M2 et M3 (dans certains sous-types également à M1) via des liens flexibles, qui limitent la distance maximale entre les domaines A et M.
2. Le domaine de phosphorylation (P) forme une insertion dans le CTM.
M4b et M5 du CTM s'étendent dans le cytosol et constituent une partie du domaine P.
Le CTM et le domaine P semblent se déplacer ensemble comme un corps semi-rigide.
3. Le domaine de liaison aux nucléotides (N), domaine N cytosolique, comprend une insertion dans le domaine P à travers une charnière fortement conservée de deux brins peptidiques antiparallèles, pointant ainsi à l'opposé de la membrane.
Sous-unités β et γ
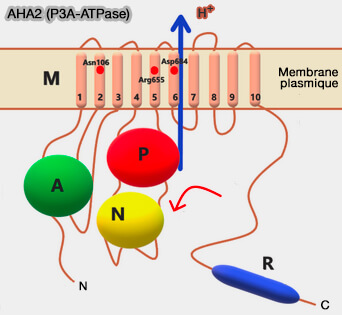
La sous-unité α peut être associée à des sous-unités supplémentaires :
L'extension N-terminale peut fonctionner comme un capteur de pH,
L'extension de la partie C-terminale ou domaine régulateur (R) est auto-inhibitrice et peut être libérée par phosphorylation ou lors de la liaison à des facteurs de régulation.





 structure des P4-ATpases ou flippases).
structure des P4-ATpases ou flippases).