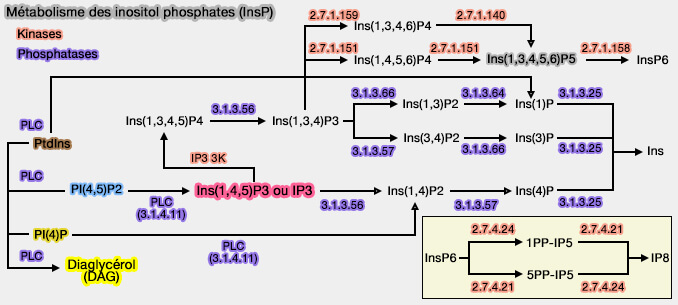
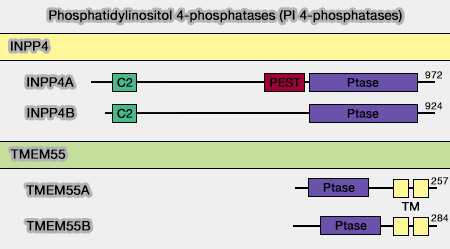
3. En outre, INPP4A contient une séquence PEST,région riches en proline (P), en acide glutamique (E), en sérine (S) et en thréonine (P), intermédiaire entre les deux domaines précédents, domaine qui permet à la calpaïne de cliver INNP4A.
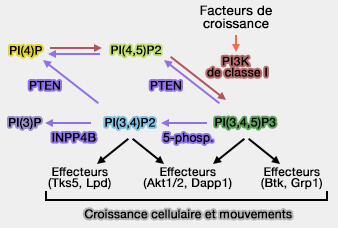
INNP4, par l'hydrolyse de PI(3,4)P2, régule localement le facteur de croissance et les signaux nutritifs au niveau des endosomes des cellules cancéreuses.
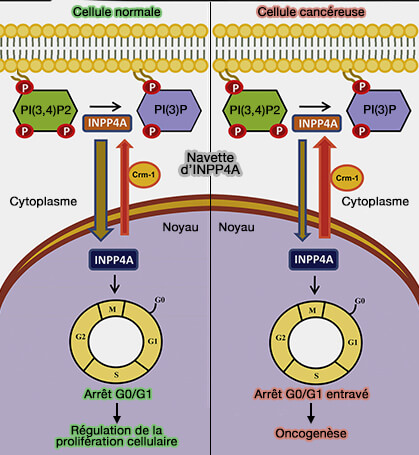
Navette d'INPP4A
(Figure : vetopsy.fr d'après Chaudhuri et coll)La synthèse et le renouvellement de PI(3,4)P2 seraient spatialement séparés au sein de la voie endocytaire pour coupler le trafic membranaireendocytaire au facteur de croissance et à la signalisation des nutriments.
L'INPP4A nucléaire est enzymatiquement actif et en équilibre dynamique entre le noyau et le cytoplasme en fonction du stade du cycle cellulaire, les quantités les plus élevées étant détectées dans le noyau pendant la phase G0/G1.
De plus, l'INPP4A nucléaire s'avère avoir une activité de suppression directe de la prolifération. Les cellules surexprimant constitutivement l'INPP4A nucléaire présentent une apoptose massive, celles la sousexprimant provoque la croissance cancéreuse.
Paradoxalement, INPP4B est également un oncogène signalé dans d'autres cancers.
Les cancers du sein ER+ mutants PIK3CA présentent une expression accrue de l'ARNm et des protéines d'INPP4B, INPP4B a augmenté la prolifération et la croissance tumorale des cellules cancéreuses du sein ER+ mutantes PIK3CA, malgré la suppression de la signalisation AKT.
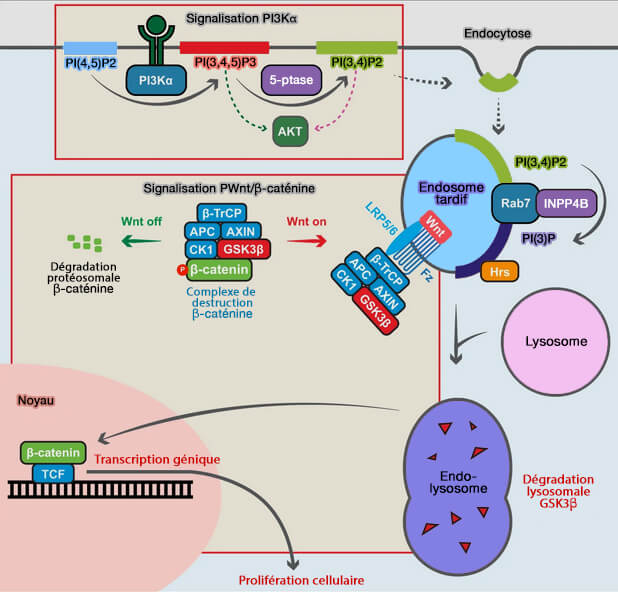
INPP4B facilite la diaphonie PI3Kα avec la signalisation Wnt dans le cancer du sein ER+ via la conversion PI(3,4)P2 en PI(3)P sur les endosomes tardifs, suggérant que ces tumeurs peuvent être ciblées avec des thérapies combinées PI3K et Wnt/β-caténine.
Signalisation AKT et Wnt dans les cancers du sein
(Figure : vetopsy.fr d'après Rodgers et coll)
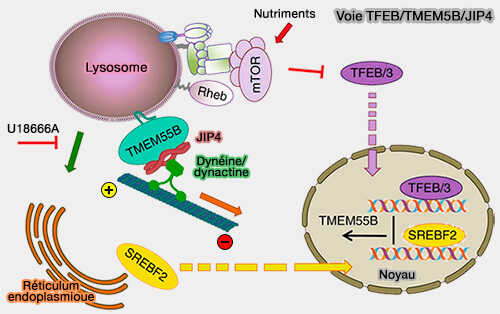
Voie TFEB/TMEM5B/JIP4
(Figure : vetopsy.fr d'après Willet et coll)
1. La voie TFEB/TMEM55B/JIP4 coordonne le mouvement des lysosomes en réponse à une variété de conditions de stress.
TMEM55B recrute JIP4 à la surface lysosomale, induisant le transport dépendant de la dynéine des lysosomes vers l'extrémité négative des microtubules.
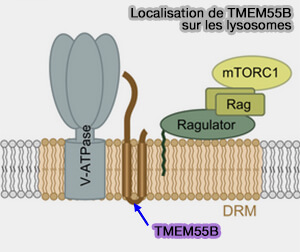
TMEM55B et lysosomes
(Figure : vetopsy.fr d'après Hashimoto)
La surexpression de TMEM55B provoque l'effondrement des lysosomes dans le centre cellulaire, tandis que l'épuisement de TMEM55B ou de JIP4 entraîne une dispersion vers la périphérie cellulaire.
Les niveaux de TMEM55B sont régulés à la hausse suite à l'activation de TFEB et TFE3 par la famine ou le stress lysosomal induit par le cholestérol. La déplétion en TMEM55B ou JIP4 abolit le transport lysosomal rétrograde induit par la famine et empêche la fusion autophagosome-lysosome.