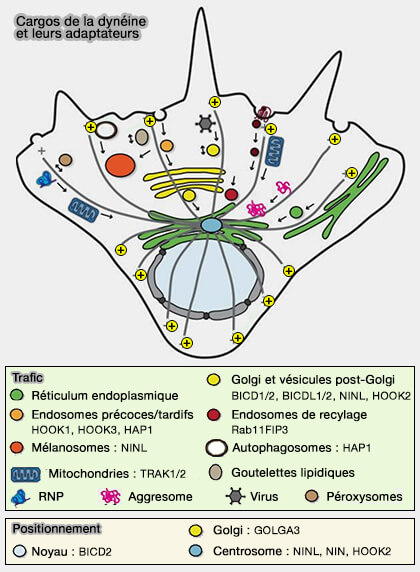
un site de liaison pour les protéines qui relient les adaptateurs aux cargos, i.e. petites GTPases Rab comme Rab6, Rab11, RZZ, FTS…
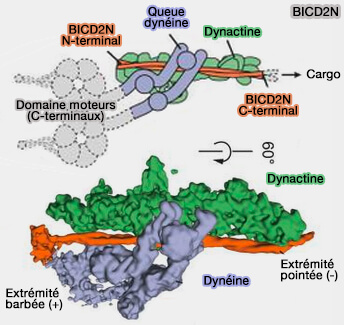
2. On peut prendre comme exemple Bicaudal-D (BicD), une protéine de 820 résidus, i.e. petite molécule comparée à la dynactine ou à la dynéine, protéine enroulée associée au transport de vésicules, d'ARNm et de noyaux.
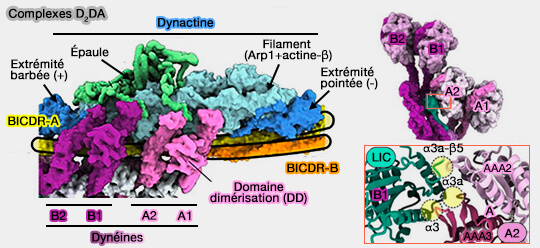
2. En outre, le domaine coiled-coil (superhélice) semble jouer un rôle sur le nombre de molécules de dynéine qui sont recrutées sur l'échafaudage de la dynactine, i.e. formation de complexes DDA ou D2DA.
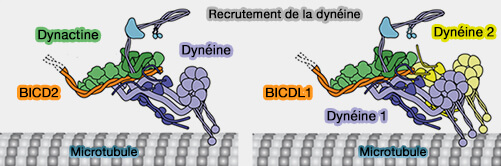
3. Certains adaptateurs, comme BICDR1, sont présents en double dans les complexes D2DA, i.e. contenant deux dimères de dynéines, ce qui permet de préciser les interactions entre les dynéines (complexes D2DA).
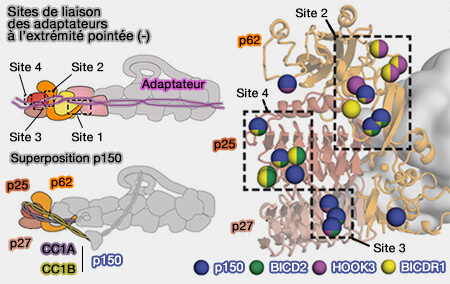
Interactions à l'extrémité pointée de la dynactine
Les interactions des adaptateurs à l'extrémité pointée de la dynactine, qui semblent moduler l'affinité entre la dynéine et la dynactine, diffèrent selon les adaptateurs à l'extrémité pointée, i.e. ils possèdent des motifs différents d'activation.
Les adaptateurs de la dynéine BICDR1, BICD2 et Hook3 utilisent des sites qui se chevauchent, mais diffèrent le long du filament de dynactine.
1. La majorité des interactions à l'extrémité pointée (-) se regroupent autour de quatre sites.
Le site 1 implique la boucle désordonnée suivant la longue hélice dans p62.
Il est partagé par les trois adaptateurs, la boucle semblant adopter différentes conformations pour lier chaque adaptateur.
Sites de liaison des adaptateurs à l'extrémité pointée (-)
(Figure : vetopsy.fr d'après Lee et coll)
Les autres sites sont contactés par différents sous-ensembles d'adaptateurs.
Le site 2 est dans la région de la selle de p62 près de p25.
Le site 3 se trouve dans une boucle qui s'étend à partir de p25.
Le site 4 se trouve sur la face d'extrémité du pli β-hélicoïdal de p25.
2. Les domaines CC1A/B du bras de p150, i.e. son extrémité N terminale, interagissent avec un ensemble distinct de résidus, qui chevauchent les sites utilisés par les adaptateurs, i.e. lorsque le bras de p150 est lié à l'extrémité pointée, les trois adaptateurs (BICD2, BICDR1 et Hook3) sont stériquement empêchés de se lier à la dynactine.
CC1A interagit avec le site 3 et le site 4.
CC1B couvre le site 2.
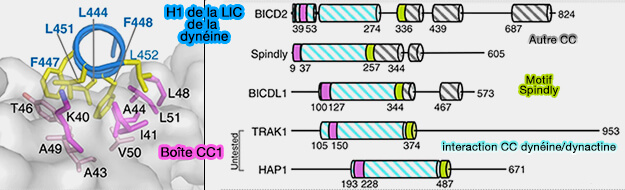
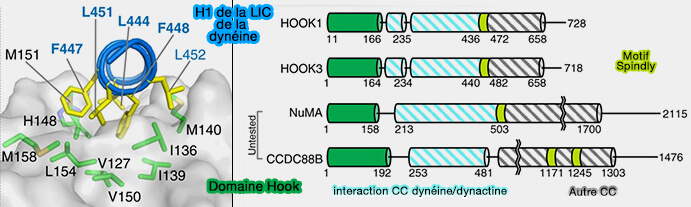
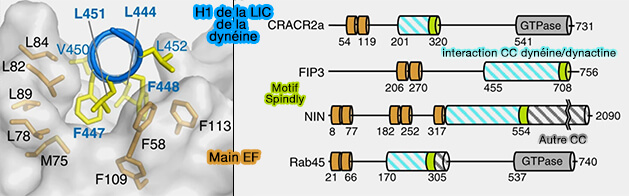
Interactions avec la chaîne légère intermédiaire (LIC) de la dynéine
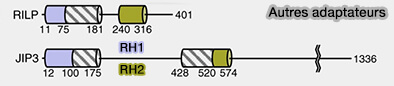
Autres adaptateurs de la dynéine
(Figure : vetopsy.fr d'après Lee et coll)
Remarque : les membres de la famille RILP et JIP partagent certaines caractéristiques avec les adaptateurs dynéine-dynactine, notamment la capacité de se lier à LIC, mais il est peu probable qu'ils activent la motilité processive.
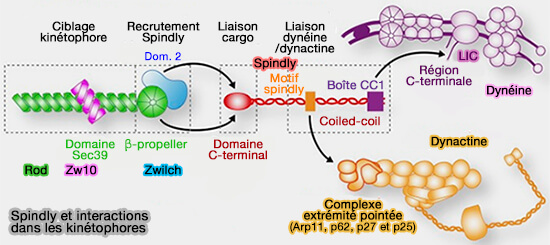
Interactions avec l'extrémité pointée (-) de la dynactine
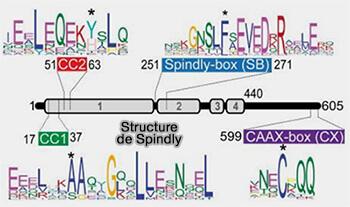
Structure de Spindly
(Figure : vetopsy.fr d'après Sacritan et coll)
Cette interaction au motif Spindly est celle qui diffère le plus selon les adaptateurs car elle est directement ou indirectement responsable du recrutement de cargos spécifiques.
Le recrutement de la sous-unité p150 de la dynactine aux kinétochores a été compromis dans les cellules exprimant GFP-Spindly muté dans l'un des trois motifs [A24V (SpindlyΔCC1), Y60A (SpindlyΔCC2), F258A (SpindlyΔSB)]et a été presque aboli lorsque les trois motifs ont été mutés (SpindlyΔCCS).




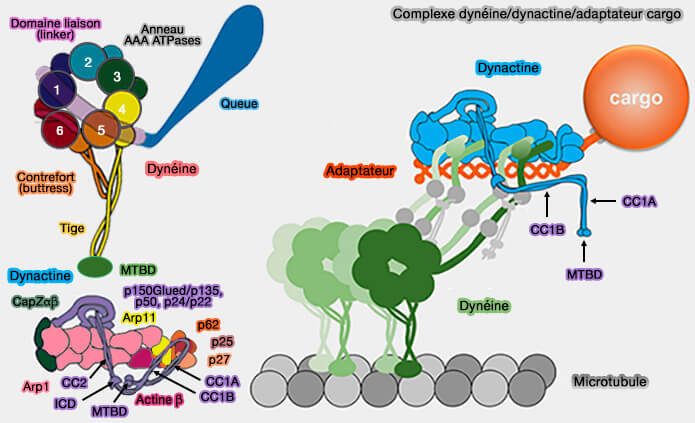
 tableau).
tableau).