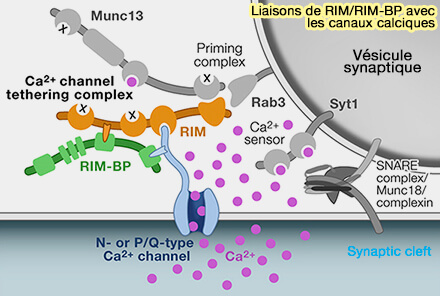
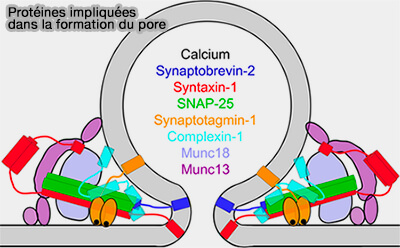
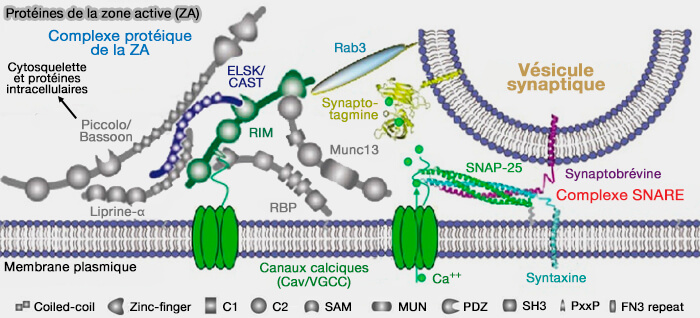
Les molécules impliquées dans les processus du cycle des vésicules synaptiques (VS) sont nombreuses et pas toutes découvertes, ce qui laisse encore envisager des révisions du modèle classique.
Protéines impliquées dans la formation du pore
(Figure : vetopsy.fr d'après Sharma et coll)
la dynamine, qui est maintenant connue pour intervenir dans la préparation de la courbure, la formation des vésicules et leur fission,
2. Ces transformations offrent de nombreux mécanismes constants ou sous-estimés pour contrôler l'exo-endocytose.
Les pores de fusion sont dynamiques et peuvent
:
se dilater jusqu'à la largeur de la vésicule,
puis, se rétrécir et se fermer, tandis que la taille du profil vésiculaire fusionné peut rester inchangée, s'agrandir, se rétrécir partiellement ou se rétrécir complètement.
Dynamique des pores de fusion
(Figure : vetopsy.fr d'après Wu et Chan)
Le kiss-and-run est redéfini pour inclure toute taille de pores qui peut limiter ou favoriser la libération et la formation de différentes tailles de vésicules selon le profil agrandi-fermé (enlarge-close) ou rétréci-fermé (shrink-close).
Les fusions par rétrécissement et élargie, plutôt que les effondrements complets et kiss-and-run classiques, utilisent les grands et les petits pores pour promouvoir et limiter la libération de contenu, respectivement.
2. Le modèle décrit doit être amélioré, car de nombreuses questions n'ont pas encore de réponses.
Comment la dynamine concurrence les protéines SNARE pour antagoniser l'hémifusion en fusion complète et l'expansion du pore ?
Comment les sites de libération sont rapidement assemblés en des profils Ω fusionnés pour supporter la fusion séquentielle ?
Comment l'actine agit avec la dynamine pour attirer la membrane à l'intérieur ?
Comment la dynamine contracte la base de grands profils Λ et les pores des profils Ω ?
Comment les protéines SNARE ouvrent le pore de fusion ?
Comment la dynamine contribue à l'hémi-fission, puis au passage de l'hémifission à la fission complète ?
Comment la clathrine et d'autres protéines de revêtement membranaire contribuent à la formation de profil Ω ?
Comment l'influx de calcium déclenche chaque transformation de la membrane exo-endocytaire ?
Comment des dizaines de protéines exo-endocytaires et de lipides contribuent à chaque transformation de la membrane exo-endocytaire ?
Comment les principes résumés ici s'appliquent à différents types cellulaires ou organites ?
Il reste du travail sur la planche pour enfin élucider tous les phénomènes complexes se déroulant dans la membrane présynaptique.



{kind=link}

 senseurs calciques et cycle vésiculaire) :
senseurs calciques et cycle vésiculaire) :