L'augmentation de la concentration en ions calcium (Ca++) a un rôle majeur dans l'électrosécrétion des vésicules synaptiques (VS).
Vue d'ensemble de l'électrosécrétion
L'électrosécrétion est la libération d'une petite quantité de neurotransmetteur contenue dans l'extrémité de l'élément synaptique suite à l'arrivée du potentiel d'action, i.e. c'est un processus électrique qui déclenche la " sécrétion " du médiateur.
Ces canaux à haut seuil ne s'ouvrent qu'en réponse à des fortes dépolarisations.
a. La concentration en Ca++ est nettement plus élevée dans le milieu extracellulaire (1-2 mmoles/L) que dans le milieu intracellulaire (10-4 mmoles/L).
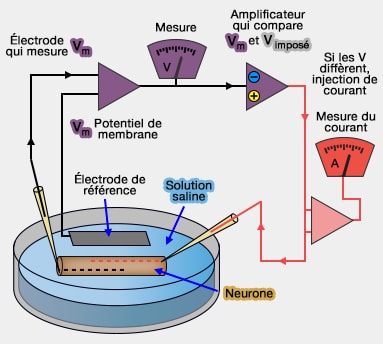
Voltage clamp
(Figure : vetopsy.fr)
b. Contrairement aux ions Na+, l'entrée de Ca++ déclenchée par le potentiel d'action entraîne une augmentation importante de sa concentration intracellulaire qui est très faible au repos.
c. Dans une expérience sur le neurone de calmar, l'injection d'un courant adéquat provoque un plateau de dépolarisation de + 20 mV. Les ions Ca++ sont soumis à deux forces ( techniques de mesure des potentiels membranaires) :
la force de diffusion, i.e. la concentration en Ca++ étant beaucoup plus élevée à l'extérieur qu'à l'intérieur de la fibre : la valeur du potentiel d'équilibre du Ca++ (VCa) est de + 130 mV. Dès lors, lorsque la dépolarisation a ouvert les électrorécepteurs Ca++, les ions Ca++, ont tendance à entrer sous l'influence de la force de diffusion.
la différence de potentiel membranaire, i.e. le potentiel de membrane de + 20 mV fait que le milieu intracellulaire est positif et s'oppose donc à l'entrée des ions Ca++. La force de diffusion (VCa = + 130 mV) est supérieure à la différence de potentiel imposée à la membrane (Vm = + 20 mV) et les ions Ca++ entrent, déclenchent une libération de médiateur et, après le classique délai synaptique, un potentiel postsynaptique est enregistré.
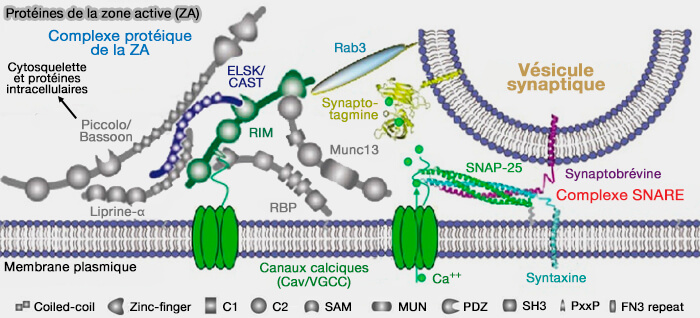
Calcium et transmission synaptique
(Figure : vetopsy.fr d'après Südhof)
3. Lors de la dépolarisation membranaire, les électrorécepteurs calcium s'ouvrent, i.e. laissent traverser sélectivement les ions Ca++ qui vont être détectés par les senseurs calciques ( senseurs calciques et électrosécrétion) comme :
Par exemple, Munc13 est particulièrement bien adaptée pour contrôler la transition LS ➞ TS, i.e. par l'interaction de son domaine C2B avec Ca++ ( processus engagés).
Lors du potentiel d'action (PA), l'afflux de calcium augmente d'un facteur 105 la vitesse d'exocytose.
5. Cette exocytose s'effectue dans un délai court, d'environ deux cents ms, parce que l'ensemble des canaux calcium, des protéines sensibles aux ions calcium et des vésicules amarrées sont très proches, i.e. dans la zone active (ZA) et liés les uns aux autres.
Protéines de la zone active (ZA)
(Figure : vetopsy.fr d'après Mochida et Wang)
Libération des neurotransmetteurs et concentration calcique
Lorsque l'entrée du calcium dépendant de la tension est supposée suffisamment importante, la sortie synaptique atteint une valeur maximale d'une vésicule synaptique par potentiel d'action et par site.
L'ajustement des résultats avec des modèles actuels d'exocytose dépendante du calcium indique que la limite de vésicule de 0,22 reflète la probabilité que les sites d'accueil soient occupés par des vésicules synaptiques au repos, car seules les vésicules amarrées peuvent être libérées.
Avec 3 mm de calcium externe, la sortie maximale par site augmente à 0,47, indiquant une augmentation de l'occupation d'accueil du site en fonction de la concentration de calcium externe.
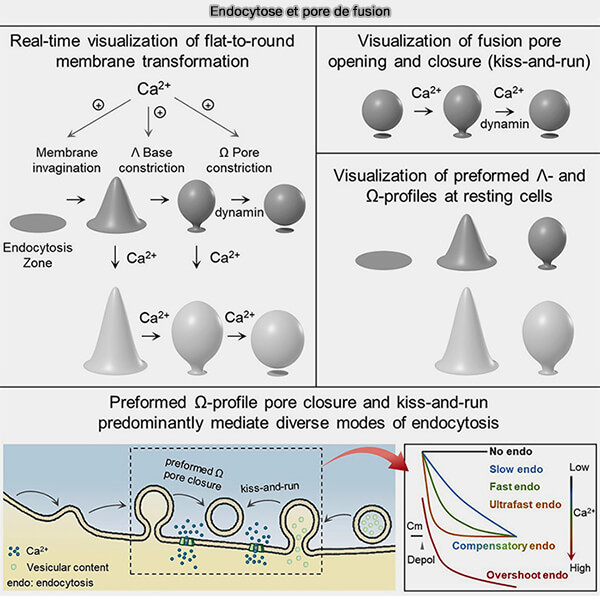
Dynamique des pores de fusion
(Figure : vetopsy.fr d'après Wu et Chan)
Retour au repos
L'augmentation du calcium intracellulaire est transitoire et brève, et le retour à l'état de repos des terminaisons axonales implique plusieurs processus.
1. Après l'excitation, la membrane va se repolariser par l'inactivation :
Les canaux KCa font sortir le potassium pour aider à repolariser la membrane.
Les ions Ca++ qui entrent dans la terminaison présynaptique activent la partie intracellulaire de ces canaux, qui s'ouvrent et laissent sortir des ions potassium.
Il en résulte une repolarisation rapide de la terminaison présynaptique par la fermeture des canaux calcium qui inhibe l'entrée des ions Ca++.
2. Les ions Ca++ sont expulsés de la région présynaptique par plusieurs processus :
des pompes, qui utilisent de l'énergie sous forme d'ATP et qui mettent le calcium soit en réserve, par exemple dans le réticulum endoplasmique, soit vers l'extérieur,
des transporteurs qui transportent, qui échangent des ions calcium contre des ions sodium et qui expulsent là encore le calcium à l'extérieur,
la liaison avec des protéines qui sont dans le milieu intracellulaire, comme la parvalbumine, la calbindine, qui sont capables de lier les ions calcium et font disparaître le calcium sous sa forme ionique, i.e. active.
Ca++ et endocytose des vésicules synaptiques
Les ions Ca++ interviennent aussi dans l'endocytose des vésicules synaptiques et déclenchent, selon leur concentration, les différents types d'endocytose ( différentes endocytoses lors du cycle synaptique).
Endocytose et pores de fusion (Figure : vetopsy.fr d'après Shin et coll)



{kind=link}

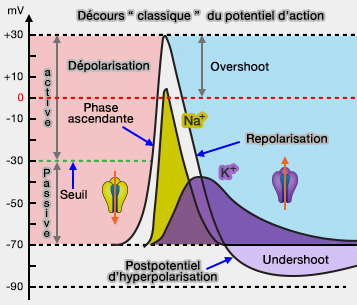
 potentiels membranaires et concentrations ioniques).
potentiels membranaires et concentrations ioniques).