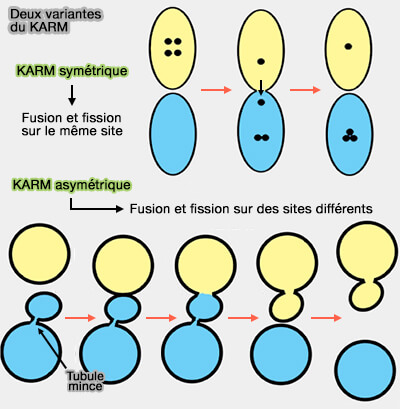
Le modèle kiss-and-run est un processus dans lequel une vésicule s'ancre et fusionne de manière transitoire au niveau d'une membrane, libère son contenu, puis se détache pour être réutilisée rapidement.
Deux vésicules entrent de manière transitoire en contact par leur membrane (kiss 1).
Deux variantes du KARM
(Figure : vetopsy.fr d'après Mironov et Beznoussenko)
Un petit pore de fusion permet un échange unidirectionnel ou bidirectionnel de molécules (kiss 2).
Les vésicules se séparent rapidement par fermeture du pore de fusion pour empêcher leur coalescence en un seul compartiment.
b. La variante asymétrique du KARM présuppose que la fusion/fission peut se produire sur des sites différents, appelé aussi modèle de maturation des transporteurs (CMM ou carrier-maturation model).
Le modèle kiss-and-run (KARM) est de plus en plus incriminé dans le trafic vésiculaire, que ce soit dans l'endocytose rapide ou dans la voie sécrétoire, et est développé dans des chapitres spéciaux ( KARM de l'EGT et KARM de l'IGT).
Fusion séquentielle, fusion composée et kiss-and-run
(Figure : vetopsy.fr d'après Wu et Chan)
Principe de base du modèle kiss-and-run (KAR)
L'endocytose est essentielle à la vie d'une cellule.
1. Dans les cellules non excitables, l'endocytose, y compris l'endocytose clathrine-dépendante (CME), prend généralement quelques minutes pour absorber les molécules de signalisation, i.e. ligands extracellulaires, récepteurs, virus, bactéries…
Le bourgeonnement dépendant de la clathrine fournit un mécanisme permettant de générer une vésicule enrichie en un cargo spécifique.
La nécessité d'un mécanisme de tri moléculaire long pour sélectionner le cargo pourrait être contournée si les vésicules ne subissent pas un effondrement complet (full-collapse fusion) dans la membrane plasmique après la fusion, conservant ainsi leur identité, ce qui est le cas pour le kiss-and-run.
2. Dans les systèmes nerveux et endocrinien, où l'exocytose peut rapidement épuiser les vésicules chargées de neurotransmetteurs/hormones, l'endocytose peut prendre seulement quelques millisecondes à des dizaines de secondes pour récupérer les vésicules en fusion (Synaptic Vesicle Endocytosis in Different Model Systems 2018).
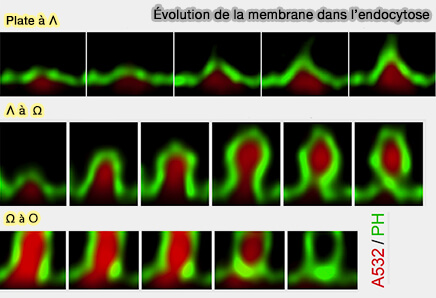
Formes des vésicules
(Figure : vetopsy.fr d'après Shin et coll)
Pour répondre à ces demandes dans des conditions physiologiques hautement dynamiques, les cellules excitables comme les neurones et les cellules endocrines développent de multiples modes endocytaires ( différents modes d'endocytose).
a. L'endocytose classique est constituée de transitions modulaires, i.e. ( profils vésiculaires) :
des vésicules plates à Λ (Plat → Λ), invagination de la membrane plate,
de vésicules Λ à Ω (Λ → Ω), constriction de la base de Λ
de vésicules Ω à O (Ω → O), constriction du pore de Ω
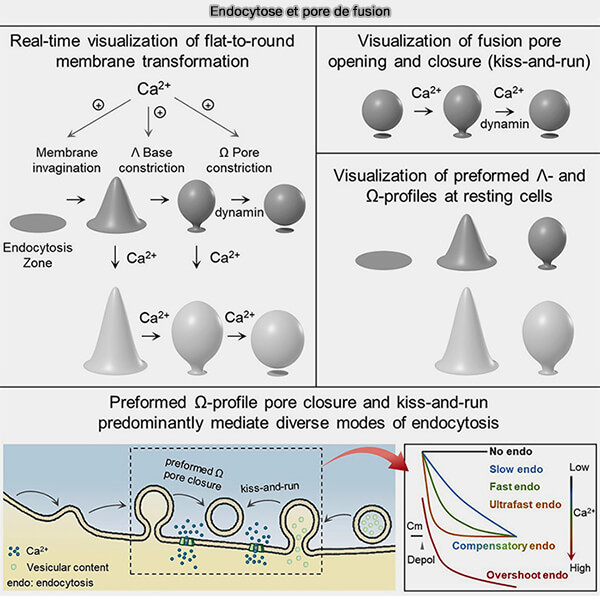
b. En outre, la formation de vésicules endocytaires résulte principalement non pas d'une transformation de membrane plate en vésicule ronde, mais d'une fermeture déclenchée par le calcium et dépendante de la dynamine ( transition et constriction) :
des pores de profils Ω préformés (pré-Ω), i.e. formés avant la dépolarisation,
des pores de fusion, i.e. kiss-and-run.
Endocytose et pores de fusion
(Figure : vetopsy.fr d'après Shin et coll)
4. Des influx de calcium variables de faibles à élevés augmentent :
a. la vitesse de fermeture des pores pré-Ω et de fusion des pores, générant une endocytose :





 KARM de l'EGT et
KARM de l'EGT et