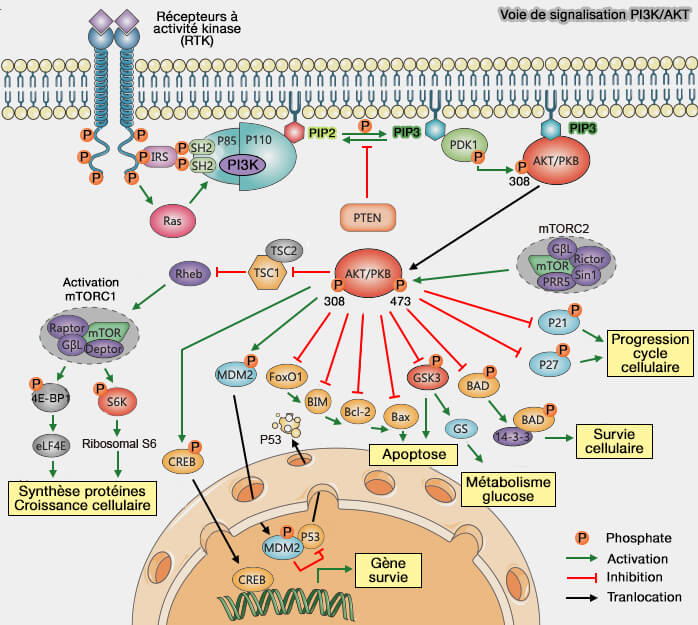
La voie PI3K/AKT est une cascade de signalisation clé qui relie les stimuli extracellulaires à la survie, la croissance, le métabolisme et la prolifération cellulaire.
3. AKT activée se transloque dans le cytoplasme et le noyau, où elle phosphoryle de nombreux substrats impliqués dans la survie, la prolifération et le métabolisme.
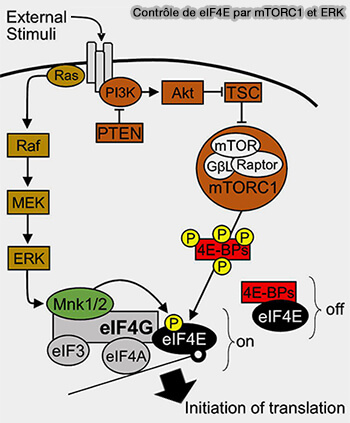
2. Une fois activé, mTORC1 phosphoryle deux cibles clés.
a. La phosphorylation de S6K1 (ribosomal protein S6 kinase 1) augmente la capacité globale de traduction en activant la protéine ribosomique S6.
S6 est une protéine constitutive du ribosome 40S impliquée dans la traduction des ARNm riches en motifs 5′TOP (Terminal Oligopyrimidine Tract), i.e. ARNm codant pour des protéines ribosomiques et des facteurs de traduction, favorisant ainsi la production des composants du ribosome et des protéines nécessaires à la croissance cellulaire.
S6K1 peut phosphoryler des substrats en amont, notamment IRS1, ce qui réduit la signalisation PI3K/Akt et constitue un mécanisme de régulation fine destiné à éviter une activation excessive du métabolisme anabolique.
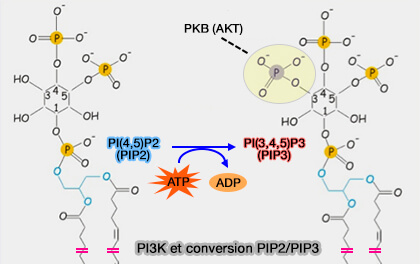
PI3K et conversion PIP2/PIP3
(Figure : vetopsy.fr d'après Amorim et coll)
eIF4E, un facteur d’initiation essentiel de la traduction des ARNm riches en structures secondaires dans leur région 5′UTR (5' Untranslated Transcribed Region) qui se trouve directement en amont du codon d'initiation.
Ainsi, l'activation de mTOR inhibe l'activité inhibitrice du complexe 4E-BP1/eIF4E.
mTORC1 phosphoryle donc les protéines en aval pour induire la croissance cellulaire et la synthèse des protéines tout en inhibant l'autophagie.
Prolifération
AKT phosphoryle p21 et p27, les inhibiteurs de CDK (Cyclin-dependent kinase), pour favoriser l’entrée en phase G1 de l'interphase en activant :
la production de la cycline D1,
les complexes cycline D/CDK4-6.
Migration et adhésion cellulaire
L’activation d’AKT influence directement la morphologie et la motilité cellulaires par plusieurs mécanismes.
Il régule l’adhésion, la polarité cellulaire et la signalisation mécano-sensible.
3. AKT contrôle l’assemblage des complexes d’adhésion focale, qui stabilisent la cellule sur la matrice et transmettent des signaux de survie et de croissance.
Cette régulation est cruciale pour la migration dirigée, l’invasion tumorale et l’adaptation à l’environnement tissulaire.
Régulation de la voie PI3K/AKT
Régulation et rétrocontrôle
1. PTEN (phosphatase and tensin homolog located on chromosome TEN) est une phosphatase qui déphosphoryle PIP3 en PIP2, inhibant la voie de signalisation PI3K/AKT.
a. Cette coactivation permet une synergie entre croissance et prolifération.
PI3K/AKT favorise la survie et le métabolisme,
MAPK/ERK stimule la progression du cycle cellulaire par l’induction de la cycline D1.
b. On trouve même des boucles de rétroaction.
AKT peut, par phosphorylation, inhiber Raf (MAPKKK), régulateur en amont d’ERK, modulant ainsi la durée et l’intensité du signal mitogène.
2. La voie PI3K/AKT exerce une influence antagoniste sur les voies des kinases JNK (c-Jun N-terminal kinase) et p38, autres familles des MAPK, activées en réponse à des stress cellulaires, i.e. oxydatif, génotoxique, ou inflammatoire.
Inversement, une activation prolongée de p38 ou JNK peut inhiber AKT via des phosphatases comme PP2A ou PTEN, orientant la cellule vers l’apoptose.
3. Ce crosstalk assure une intégration hiérarchique des signaux.
Les voies MAPK contrôlent la prolifération et la différenciation.
La voie PI3K/AKT préserve la viabilité cellulaire et le métabolisme énergétique.
Dans de nombreux contextes, notamment tumoraux, la coactivation persistante de ces voies renforce la résistance aux signaux apoptotiques et favorise la progression néoplasique.
Implications pathologiques
1. Les implications pathologiques sont nombreuses.
a. L'hyperactivation de la voie PIK3/AKT provoque :
des cancers,
une résistance aux thérapies,
des maladies métaboliques.
b. L'hypoactivation entraîne :
des troubles du développement,
des déficits de survie cellulaire.
Remarque : cette activation est souvent la conséquence de mutations de PTEN, de la sous-unité catalytique p110α de PI3K, la rendent constitutivement active, ou d'AKT.
2. Les Inhibiteurs ciblés de PI3K, AKT et mTOR pourraient moduler cette voie pour réguler l'apoptose, le métabolisme et la prolifération.