Les ligases CRL utilisent des adaptateurs modulaires pour relier la culline à leurs récepteurs de substrats, assurant la spécificité et la diversité fonctionnelle du système ubiquitine-protéasome.
Vue d'ensemble des adaptateurs des CRL
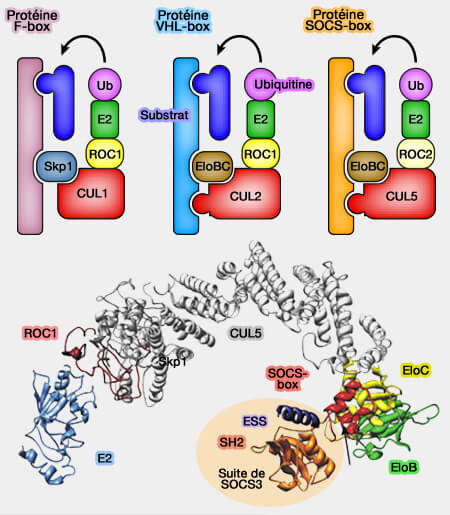
Les ligases CRL (Cullin-RING ligases) représentent la principale sous-famille des ligases E3 de type RING qui se distinguent par leur architecture modulaire où la culline agit comme échafaudage central reliant ( organisation modulaire des CRL des CRL) :
un récepteur de substrat conférant la spécificité du complexe.
Ces adaptateurs, souvent spécifiques d’un sous-type de culline, servent de ponts structuraux essentiels entre la culline et la protéine réceptrice, permettant à chaque type de complexe CRL de cibler une vaste gamme de substrats, tout en conservant une architecture catalytique commune.
Remarque : les ligases RING monomériques, telles que MDM2 ou c-Cbl, reconnaissent directement leurs substrats sans adaptateur intermédiaire.
Principales familles d’adaptateurs des complexes CRL
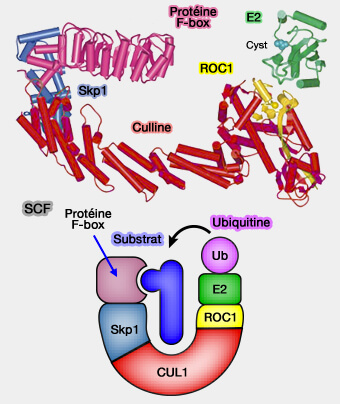
Adaptateurs Skp1 et protéines F-box (complexe SCF/CUL1)
1. Skp1 (S-phase kinase-associated protein 1) est l’adaptateur canonique de CUL1, reliant la culline à la protéine F-box.
SCF
(Figure : vetopsy.fr)
Sa structure compacte, de 163 résidus, formée de feuillets β et d’hélices α, crée une surface hydrophobe stable qui s’emboîte dans le domaine F-box, stabilisant ainsi l’interface avec la culline.
2. Dans le complexe SCF (Skp1-Cullin1-F-box-Rbx1), pivot de la dégradation cyclique des régulateurs du cycle cellulaire, elle assure la liaison entre la culline 1 (CUL1) et la protéine F-box en faisant partie du " fer à cheval ", responsable de la reconnaissance du substrat ( figure).
2. Les protéines à domaine DWD constituent une sous-classe majeure de DCAF.
a. Ce motif de 16-17 acides aminés, inclus dans une lame WD40, présente un consensus typique WDxR et forme un site d’interaction situé à la surface du propeller WD40, assurant la fixation stable à DDB1.
Ces adaptateurs établissent ainsi le pont fonctionnel entre DDB1 et les substrats, assurant la sélectivité des complexes CRL4.
b. Ces protéines DWD jouent donc un rôle équivalent à celui des F-box (complexes SCF) ou des BTB (complexes CUL3), mais au sein des E3 ligases CRL4, contribuant à la diversité fonctionnelle et à la spécificité des substrats du système ubiquitine-protéasome.
c. On peut citer comme exemple :
DDB2, impliquée dans la réparation de l’ADN endommagé par les UV ;
CSA (Cockayne syndrome A), intervenant dans la réponse au stress oxydatif et la réparation couplée à la transcription,
DET1, participant à la régulation de la photomorphogenèse et des signaux lumineux ;
WDR23, modulant la stabilité du facteur NRF2, acteur majeur de la réponse antioxydante ;
CDT2 (DCAF2), assurant le contrôle de la réplication en ciblant CDT1 pour la dégradation.
Rôle fonctionnel commun
Les adaptateurs modulaires des ligases CRL assurent la spécificité de substrat et la diversité fonctionnelle du système ubiquitine-protéasome.
Ils permettent :
l’intégration de signaux post-traductionnels, i.e. phosphorylation, stress, hypoxie,
la régulation dynamique via la neddylation des cullines,




 organisation modulaire des CRL des CRL) :
organisation modulaire des CRL des CRL) :