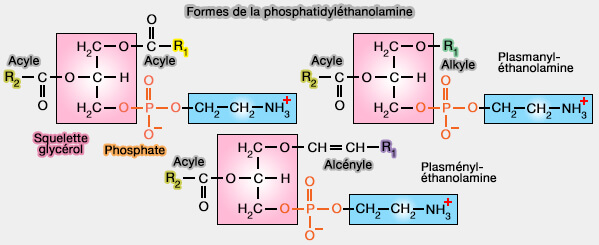
Les plamalogènes sont des glycérophospholipides qui contiennent obligatoirement une forme alcényle ($\ce{CH2-O-CH=CH-R}$) en sn-1 du squelette glycérol.
2. Deux queues hydrophobes sont formées chacune par un acide gras.
a. La liaison ester ($\ce{CH2-O-C(=O)-R}$ au sn-2 du glycérol est classique.
b. La liaison éther, dans ce cas alcényle éther (alkényl éther ou encore vinyl éther), i.e. $\ce{CH2-O-CH=CH-R}$) au sn-1 est la caractéristique des plasmalogènes, i.e. forme plasményle.
Remarque : bien qu'elles fassent aussi partie des étherlipides, la forme alkyle éther, i.e. $\ce{CH2-O-R}$, appelée plasményle, ne fait pas partie des plasmalogènes.
Formes de la phosphatidyléthanolamine (PE)
(Figure : vetopsy.fr)
3. Les acide gras liés au glycérol sont :
en sn-1 attaché à l'éther vinylique sont le plus souvent en C16:0, C18:0 ou C18:1,
Les plasmalogènes sont classés en fonction de leur groupe de tête, principalement en :
plasmalogènes PE (plasményléthalomines),
plasmalogènes PC (plasménylcholines).
1. En dehors du cœur, la plasményléthanolamine a tendance à être le principal étherlipide :
jusqu'à 80% du total des éthanolamines-phospholipides dans certains tissus cérébraux
20-40 % dans les reins, les muscles squelettiques et la rétine.
2. Beaucoup moins de phosphatidylcholine et généralement peu ou aucun des autres phospholipides, tels que le phosphatidylinositol ou la phosphatidylsérine, sont sous cette forme.
Dans le coeur, la phosphatidylcholine est en grande partie sous cette forme, i.e. dans le coeur de boeuf, 124% de diacyle, 16% d'alcényle et 1% d'alkyle.
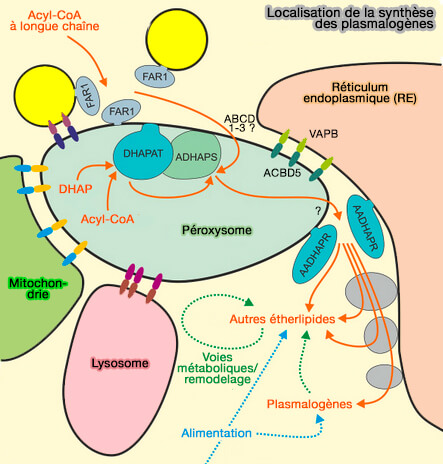
Localisation de la synthèse des plasmalogènes
Figure : vetopsy.fr d'après Farré et coll)
Dans les autres tissus, on trouve une proportion plus élevée de forme O-alkyle pour la phosphatidylcholine comme dans les neutrophiles qui en contiennent plus de 40%.
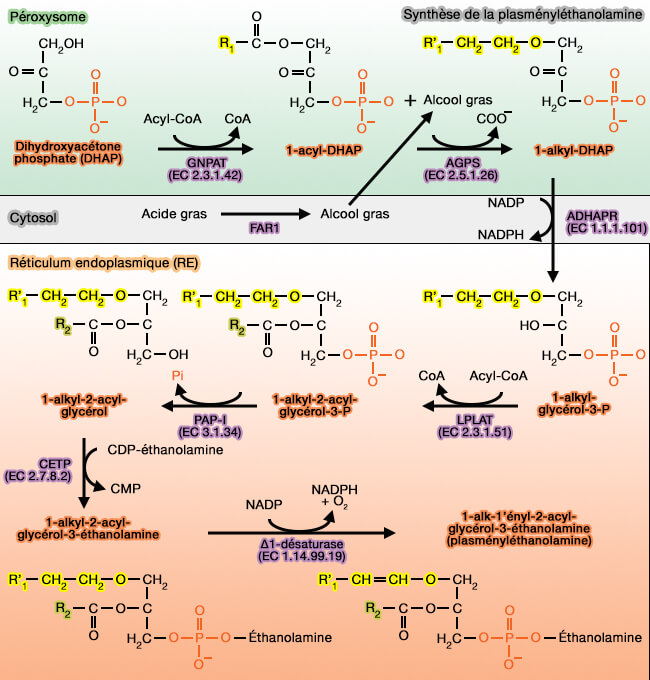
Synthèse des plasmalogènes
Contrairement aux autres phospholipides, la première étape de la synthèse se passe sur la membrane des peroxysomes.
soit produits dans le peroxysome par la β-oxydation d'esters CoA d'acides gras à très longue chaîne.
Synthèse de la plasményléthanolamine
(Figure : vetopsy.fr)
2. La deuxième étape est la formation de la liaison éther produite en échangeant le groupe acyle par un alcool à longue chaîne, réaction catalysée par AGPS (alkylglycéronephosphate synthase ou alkyl-DHAP ou ADHAPS), i.e. EC 2.5.1.26.
$\ce{1-acy-DHAP + alcool gras à longue chaine}$ $\leftrightharpoons$ $\ce{1-O-alkylDHAP + acide gras à longue chaine+ H(+)}$
a. Cette réaction est complexe et nécessite la liaison du 1-acyl-DHAP à l'AGPS, puis la réaction de l'acyl-DHAP avec FAD qui libère l'acide gras aux deux atomes d'oxygène de la liaison ester de l'acyl-DHAP, liaison de l'alcool gras à l'AGPS, et enfin formation de phosphate de 1-alkyldihydroxyacétone (1-alkyl-DHAP). L'atome d'oxygène, fait rare, provient de la fraction alcool et non du glycérol.
b. L'alcool gras à longue chaine est produite dans le cytosol par FAR1 (Fatty Acid acyl-CoA réductase 1), enzyme ancrée à la membrane peroxysomale par son extrémité C-terminale hydrophobe se lie à Pex19p, un récepteur cytosolique à motif CAAX C-terminal (Topogenesis and Homeostasis of Fatty Acyl-CoA Reductase 1 2013).
Elles débutent par la formation du 1-alkyl-G3P par l'acyl/alkyl-DHAP réductase (ADHAPR), i.e. EC 1.1.1.101, enzyme à NADP.
Les autres réactions ont lieu au niveau du RE pour obtenir la plasményléthanolamine ou 1-alk-1′-enyl-2-acyl-GPE, i.e. GPE étant la glycérol-3-phosphoéthanolamine ( cf. figure ci-dessus).
4. La plasménylcholine est obtenu à partir de la plasményléthanolamine.
La chaîne acyle sn-2 des plasmalogènes est orientée perpendiculairement à la surface de la membrane, contrairement à celle des diacylglycérophospholipides qui est courbée.
En sn-1, le groupe de tête, dépourvu d'oxygène carbonyle $\ce{C=0}$, est beaucoup plus lipophile.
1. Le plasmalogène occupe ainsi moins de surface/molécule dans un plan membranaire qu'un homologue diacyle et donc, l'interaction plus forte entre les groupes de tête a tendance à former une phase inverse hexagonale-II (formation de non-bicouche), qui est une exigence pour la fusion membranaire.
Les formes de phosphatidylcholine ont tendance à produire des phases lamellaires.
2. Les membranes cellulaires contenant des plasmalogènes sont donc moins fluides que celles déficientes en plasmalogènes, c'est-à-dire qu'elles forment des bicouches lipidiques plus comprimées, plus épaisses et rigides, propriété particulièrement importante dans les structures membranaires compactes présentes dans la myéline.
3. Malgré les concentrations relativement élevées d'acides gras polyinsaturés, les plasmalogènes ont tendance à s'accumuler dans les domaines des radeaux membranaires.
Plasmalogènes et signalisation
Les radeaux membranaires sont enrichis en cholestérol et en sphingolipides où se concentrent de nombreuses protéines de signalisation, avec des effets sur la compartimentation et les activités des enzymes membranaires.





 synthèse des triglycérides) avec un acide gras à longue chaîne par GNPAT (Glycerone-3-phosphate acyltransférase ou DHAPT), i.e. EC 2.3.1.42.
synthèse des triglycérides) avec un acide gras à longue chaîne par GNPAT (Glycerone-3-phosphate acyltransférase ou DHAPT), i.e. EC 2.3.1.42.