


Acides nucléiques
Chromatine
Chromosomes
Centromère : mécanisme global de l'incorporation de CENP-A
- Biochimie
- Chimie organique
- Bioénergétique
- Composition de la matière vivante
- Glucides
- Lipides
- Coenzymes
- Hormones
- Composés inorganiques

L’incorporation de CENP-A correspond au processus d’assemblage de la variante histonique centromérique dans la chromatine, assurant la transmission épigénétique de l’identité du centromère par des mécanismes hautement régulés impliquant HJURP et le complexe Mis18.
Le mécanisme complet de l'incorporation est complexe.
Activation du dépôt de CENP-A
L'incorporation de CENP-A est strictement limitée au début de la phase G1, première phase de l'interphase qui suit la mitose.

Vous pouvez lire : The centromere comes into focus: from CENP-A nucleosomes to kinetochore connections with the spindle (2020) qui résume les différentes étapes
1. Contrairement aux histones canoniques H3.1 et H3.2, incorporées de manière couplée à la réplication de l’ADN en phase S, le dépôt de CENP-A repose sur une succession d'événements de phosphorylation et de déphosphorylation qui contrôlent l'activation du complexe Mis18 et le recrutement de son chaperon HJURP.
a. Pendant la majeure partie du cycle cellulaire, le complexe cycline B/CDK1 (MPF) maintient la machinerie de dépôt de CENP-A dans un état inactif en phosphorylant plusieurs résidus :
- de Mis18BP1,
- du domaine R1 de HJURP (Ser412, Ser448 et Ser473).
b. À la sortie de mitose, la chute de l'activité de CDK1 entraîne leur déphosphorylation, autorisant le recrutement du complexe Mis18 puis de HJURP au niveau des centromères.
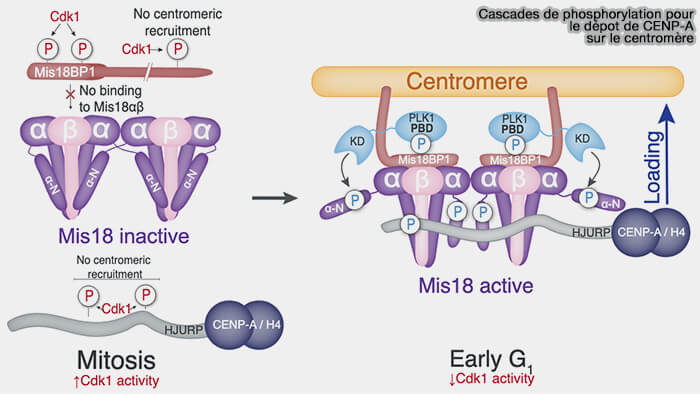
(Figure : vetopsy.fr d'après Parashara et coll)
2. Au début de G1, PLK1 (polo-like kinase 1) est recrutée au centromère de manière dépendante du complexe Mis18 et est impliquée dans plusieurs événements de phosphorylation (Role of PLK1 in the epigenetic maintenance of centromeres 2024 et PLK1-Mediated Phosphorylation Cascade Activates the Mis18 Complex to Ensure Centromere Inheritance 2024)
- PLK1 se lie à Mis18BP1 sur un site de liaison hautement conservé généré par la phosphorylation de deux résidus voisins (Thr78 et Ser93) qui doivent tous les deux être phosphorylés pour réussir le recrutement de PLK1 dans les centromères.
- Ces sites de phosphorylation sont distincts de ceux ciblés par CDK1 (Thr40 et Ser110).
3. PLK1 favorise également le recrutement de molécules supplémentaires de PLK1 par la phosphorylation de Ser54 sur MIS18α et Thr654 sur HJURP.
a. Ces phosphorylations :
- augmentent l'affinité entre HJURP et le complexe Mis18α/β,
- lèvent l'inhibition intramoléculaire du complexe Mis18,
- permettent le recrutement efficace de HJURP au centromère.
b. La perturbation du site d’amarrage de PLK1 sur Mis18BP1 (T78A et S93A) abolit le recrutement centromérique de PLK1 sans empêcher significativement la localisation du complexe Mis18.

PLK1 agit principalement comme un facteur d'activation (« licensing ») de la machinerie de dépôt de CENP-A plutôt que comme un facteur de recrutement du complexe Mis18 (Polo-like Kinase 1 Licenses CENP-A Deposition at Centromeres 2014).
Préparation de la chromatine centromérique
Après l'activation de la machinerie de dépôt, la chromatine centromérique subit plusieurs remaniements qui préparent l'incorporation de nouveaux nucléosomes contenant CENP-A.
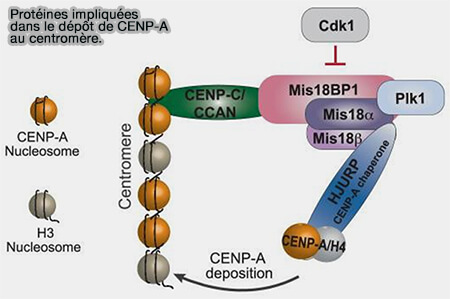
(Figure : vetopsy.fr d'après Thamkachy et coll)
1. Les lysines des queues N-terminales des histones H3 canoniques dans la chromatine centromérique sont désacétylées, ce qui ( code des histones) :
code des histones) :
- augmente l’affinité de l’ADN pour les histones,
- prépare le centromère pour un dépôt précis de CENP-A.
L’activité des histone désacétylases (HDAC) est modulée par des facteurs centromériques et par l’état épigénétique local, comme la présence de CENP-C, la configuration des satellites α…
Cette désacétylation s'inscrit dans un ensemble coordonné de signaux qui définissent le site d’incorporation de CENP-A et empêchent le dépôt aléatoire ailleurs dans le génome.
2. La chromatine est d'abord localement remodelée par des complexes de remodelage de la chromatine qui assurent l’ouverture transitoire de la structure nucléosomique.
a. RSF (Remodeling and Spacing Factor), constitué de RSF1 et de SNF2h, repositionne ou écarte localement les nucléosomes H3/H4.
- Le complexe RSF, via son activité ATP-dépendante, induit un relâchement partiel des nucléosomes H3 préexistants dans la région centromérique.
- Ce remodelage génère des fenêtres d’ADN partiellement dénucléosomisée, correspondant à des zones où l’ADN se détache temporairement du cœur histonique qui faciliteront le recrutement du complexe Mis18-HJURP et l’incorporation du tétramère (CENP-A/H4)\₂.
b. FACT (Facilitates Chromatin Transcription), bien qu’impliqué initialement dans la transcription, agit ici pour déstabiliser temporairement les interactions ADN/histones, en particulier les dimères H2A/H2B.
c. Ces complexes relâchent l’enroulement de l’ADN autour du nucléosome H3 existant ou autour de régions transitoirement dénucléosomisées.
3. L’ouverture n’est pas aléatoire, mais ciblée sur des régions marquées par CENP-C et CENP-N, qui servent de repères structuraux.
Ces protéines ancrées au CCAN (Constitutive Centromere-Associated Network) recrutent le complexe Mis18 (Mis18α/β et Mis18BP1), qui constitue la plateforme d'amorçage du dépôt de CENP-A.
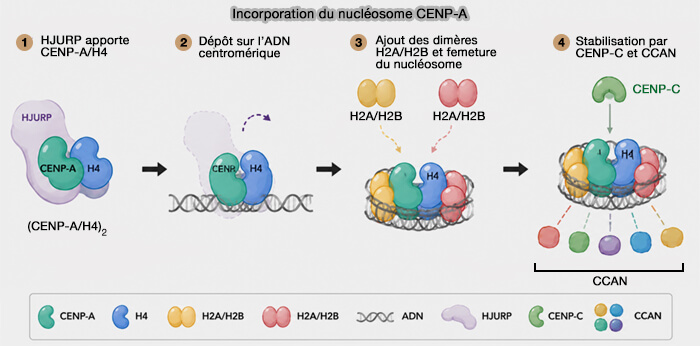
Recrutement du complexe CENP-A/H4-HJURP
HJURP (Holliday Junction Recognition Protein) forme un complexe stable avec un dimère CENP-A/H4 et le dirige vers le centromère par son interaction avec Mis18BP1 et les protéines centromériques CENP-C et CENP-N (Structure of a CENP-A–histone H4 heterodimer in complex with chaperone HJURP 2011).
1. HJURP intervient sur les fenêtres d’ADN partiellement dénucléosomisées pour positionner le dimère CENP-A/H4 sur l’ADN, sur une région d'environ 40 pb par :
- des interactions électrostatiques, i.e. résidus basiques (lysines, arginines) des queues N-terminales et de l’hélice α1 de CENP-A et d'H4 avec les sillons mineur et majeur de l’ADN.
- des interactions π-π ainsi que des interactions hydrophobes entre certaines tyrosines et arginines au niveau de CENP-A/H4 et les bases azotées, participant à la courbure et au positionnement exact de l’ADN.
(Figure : vetopsy.fr d'après Sekulic et coll)
2. HJURP :
- recouvre partiellement le dimère et interagit avec la face externe du dimère CENP-A/H4,
- protège les surfaces hydrophobes,
- empêche l’agrégation,
- maintient le dimère positionné correctement sur l’ADN.
3. Cet état est transitoire car certains segments de l’ADN restent libres ou faiblement associés, permettant l’ajout ultérieur d’H2A/H2B pour former le nucléosome complet.
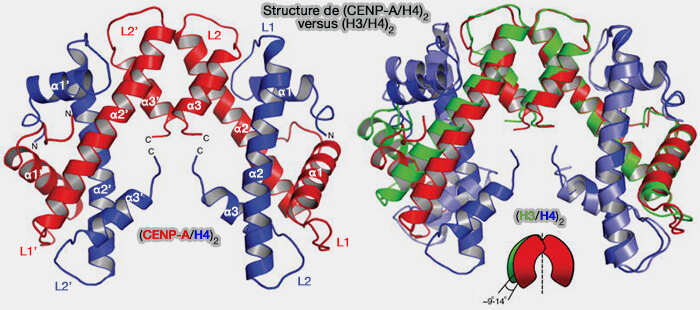
Assemblage du tétramère
Deux complexes CENP-A/H4. se positionnent successivement sur environ 80 pb d'ADN centromérique (environ 40 pb par dimère), grâce à l’action coordonnée de HJURP et du complexe Mis18α/β-Mis18BP1, qui ancre le site d’assemblage au centromère via CENP-C (Chromatin assembly: Journey to the CENter of the chromosome 2016).
1. Les interactions entre les domaines de repliement histonique des CENP-A et les queues N-terminales des H4 favorisent la dimérisation des deux dimères pour former un tétramère (CENP-A/H4)2.
a. L’interface tétramérique repose sur des interactions hydrophobes et ioniques entre les hélices α2 des domaines histoniques de CENP-A et d’H4 (
Les résidus spécifiques de CENP-A, notamment la substitution de l’alanine-87 et de la leucine-112 (comparés à H3), modifient l’angle du repli histonique, créant un tétramère plus rigide et compact.
b. La queue C-terminale de CENP-A interagit directement avec CENP-C, assurant l’ancrage de la structure au réseau du CCAN.
(Figure : vetopsy.fr)
2. HJURP reste associée au dimère CENP-A/H4 pendant cette étape pour stabiliser la structure et éviter l’agrégation ou l’incorporation erronée de CENP-A.
3. Une fois le tétramère correctement assemblé, HJURP se dissocie progressivement du complexe et est recyclée pour un nouveau cycle d'incorporation.
a. Cette dissociation est favorisée par la régulation cyclique impliquant CDK1 et Mis18BP1.
b. Parallèlement, l'établissement des interactions entre la queue C-terminale de CENP-A et CENP-C du CCAN (Constitutive Centromere-Associated Network) remplace progressivement l'ancrage transitoire assuré par HJURP.
La diminution de l'affinité du domaine SCM3 de HJURP pour CENP-A, associée aux réarrangements structuraux du complexe, favoriserait alors son détachement et son recyclage.
Remarque : les remodelases RSF et FACT de l'étape suivante facilitent le détachement.
4. Ce tétramère symétrique en tête-bêche forme le noyau central du futur nucléosome CENP-A, cœur du futur nucléosome CENP-A autour duquel s’enroule l’ADN.
Fermeture du nucléosome
Les dimères canoniques H2A/H2B non spécifiques se fixent ensuite sur les deux faces du tétramère (CENP-A/H4)2.
- Ces sites d’interaction se trouvent le long de l’axe transversal du nucléosome, à la jonction entre l’ADN d’entrée/sortie et le tétramère central.
- C’est pourquoi on dit parfois que les dimères H2A/H2B sont positionnés sur les faces latérales ou externes : ils encadrent le cœur (CENP-A/H4)2 sans en faire partie intégrante.
1. L’ADN centromérique (~121-145 pb) s’enroule maintenant complètement autour du cœur nucléosomique, stabilisant la structure et réduisant la flexibilité locale de l’ADN.
Les queues N-terminales des histones participent à la stabilisation de l’ADN par des contacts avec les sillons mineur et majeur et par des interactions électrostatiques avec le phosphate de l’ADN.
2. Les remodelases RSF et FACT interviennent pour assurer le repositionnement précis de l’ADN autour du tétramère et pour assurer que le nucléosome final soit correctement orienté.
Remarque : ils contribuent également à libérer HJURP en favorisant le remodelage des interfaces histone–ADN, facilitant son détachement une fois que le nucléosome CENP-A/H4 est correctement assemblé.
3. Le nucléosome complet est désormais structuralement stable, prêt à servir de point d’ancrage pour le réseau CCAN (Constitutive Centromere-Associated Network) et, plus tard, pour l’assemblage du kinétochore.
Il constitue un marqueur épigénétique stable du centromère, assurant la transmission de l'identité centromérique au cours des cycles cellulaires suivants.
Stabilisation du nucléosome CENP-A
Une fois assemblé, le nucléosome contenant CENP-A est stabilisé par de multiples interactions avec les histones, l'ADN et les protéines centromériques.
1. Les principales interactions structurales comprennent :
- des liaisons intranucléosiques entre CENP-A/H4 et H2A/H2B,
- des liaisons avec l’ADN et la chromatine environnants par les queues N-terminales des histones.
2. Les composants du CCAN, i.e. CENP-C, CENP-T… interagissent avec le nucléosome CENP-A pour :
- maintenir la position centromérique,
- servir de plateforme pour l’assemblage du kinétochore,
- protéger le nucléosome contre le remplacement ou la perte de CENP-A.
3. Le nucléosome CENP-A nouvellement formé est hautement stable et peut survivre jusqu’à plusieurs cycles cellulaires, assurant :
- la continuité de l’identité centromérique,
- la transmission de cette identité aux cycles cellulaires suivants grâce au dépôt de nouveaux nucléosomes contenant CENP-A au début de la phase G1 du cycle suivant.
Organisation du centromère et assemblage du kinétochore
Une fois le nucléosome contenant CENP-A stabilisé, d'autres protéines centromériques s'associent à la chromatine centromérique pour assurer son organisation et préparer l'assemblage du kinétochore.
- CENP-B se lie spécifiquement à la boîte CENP-B des répétitions α-satellites et contribue à l'organisation de la chromatine centromérique.
- Les protéines constitutives du réseau CCAN (Constitutive Centromere-Associated Network) s'assemblent ensuite autour des nucléosomes contenant CENP-A pour former la plateforme du kinétochore interne, qui servira au recrutement des autres composants du kinétochore.
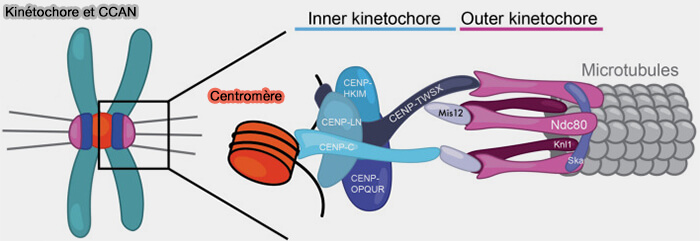
(Figure : vetopsy.fr d'après Altemose et coll)
BiochimieChimie organiqueBioénergétiqueAcides nucléiquesNucléotidesADNARNChromatineStructure dynamique de la chromatineNucléosomesHistonesMembres des histonesCode des histonesEuchromatine, hétérochromatine et remodelageChromosomesStructure des chromosomesCentromèreTélomèreProtidesGlucidesLipidesEnzymesCoenzymesVitaminesHormonesComposés inorganiquesTransport membranaireMoteurs moléculairesVoies de signalisation