


Acides nucléiques
ADN : épigénétique
Régulation
- Biochimie
- Chimie organique
- Bioénergétique
- Composition de la matière vivante
- Transport membranaire
- Moteurs moléculaires
- Voies de signalisation

La régulation épigénétique repose sur des mécanismes de mémoire et de reprogrammation qui permettent de transmettre ou de remodeler durablement les états d'expression génique sans modifier la séquence nucléotidique de l'ADN.
Les principaux mécanismes épigénétiques reposent sur :
- les marques épigénétiques, notamment méthylation de l'ADN et modifications post-traductionnelles et variants des histones,
- le remodelage de la chromatine,
- l'action de certains ARN non codants.
(Figure : vetopsy.fr)
Après leur mise en place, les marques épigénétiques ne sont pas figées et peuvent être :
- conservées au cours des divisions cellulaires afin d'assurer la stabilité des programmes d'expression génique,
- remodelées ou effacées pour permettre leur adaptation au cours du développement ou en réponse à des facteurs physiologiques ou environnementaux.
Cette capacité à concilier stabilité et plasticité repose sur des mécanismes complémentaires assurant soit la transmission, soit la reprogrammation de l'information épigénétique.
Mémoire épigénétique de la chromatine
La mémoire épigénétique est la capacité d'une cellule à conserver durablement un état d'expression génique au cours des divisions cellulaires, sans modification de la séquence nucléotidique de l'ADN.

La mémoire génétique repose principalement sur la transmission des marques épigénétiques et de certains états de la chromatine lors des divisions cellulaires.
Cette héritabilité épigénétique permet de préserver l'identité cellulaire et la stabilité des programmes d'expression génique au cours du développement et du renouvellement des tissus.
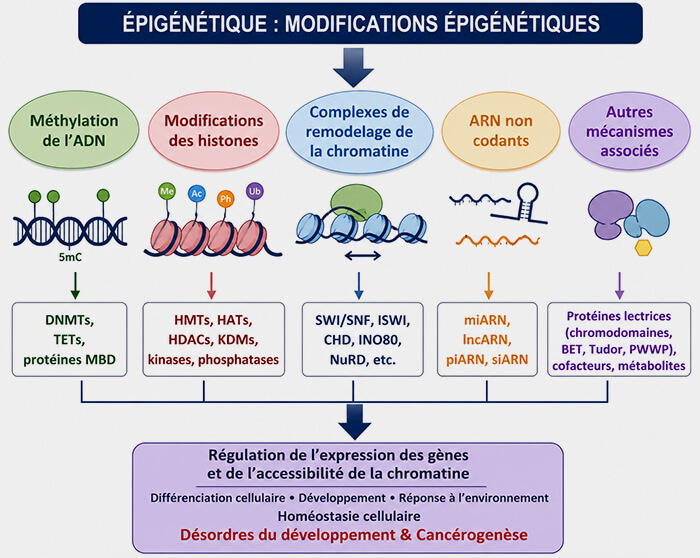
Maintien de la méthylation de l'ADN
Lors de la réplication de l'ADN, le passage de la fourche de réplication produit deux molécules d'ADN filles dans lesquelles les sites CpG sont transitoirement hémi-méthylés :
- le brin parental conserve sa méthylation,
- le brin nouvellement synthétisé en est dépourvu.
(Figure : vetopsy.fr modifiée d'après Vuu et coll)
1. La protéine UHRF1 reconnaît spécifiquement ces sites CpG hémi-méthylés et recrute la méthyltransférase DNMT1.
DNMT1 méthyle les cytosines correspondantes du nouveau brin, rétablissant ainsi le profil de méthylation initial.
2. À chaque division cellulaire, ce mécanisme permet de transmettre fidèlement les profils de méthylation de la cellule mère aux cellules filles, contribuant au maintien de leur identité, de leur programme d'expression génique et de la mémoire épigénétique.
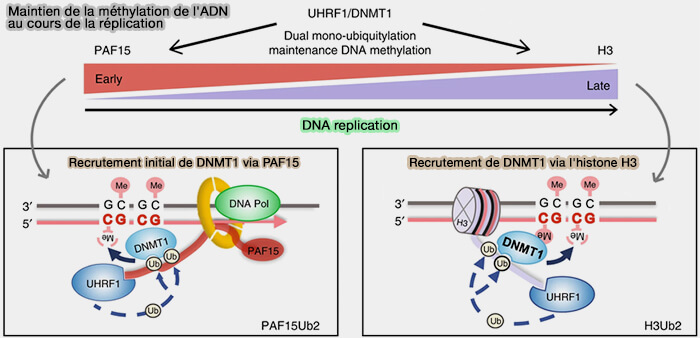
Transmission des modifications des histones
Au cours de la réplication, les nucléosomes sont temporairement désassemblés devant la fourche de réplication ( desassemblage et réassemblage des nucléosomes).
desassemblage et réassemblage des nucléosomes).
- Une partie des histones parentales, porteuses de modifications épigénétiques, est ensuite redistribuée sur les deux chromatides sœurs (Chromatin structure and the inheritance of epigenetic information 2010).
- De nouvelles histones sont simultanément incorporées afin de reconstituer les nucléosomes.
1. Les modifications présentes sur les histones parentales servent alors de modèle au recrutement de protéines readers et de complexes enzymatiques writers, capables d'établir les mêmes modifications sur les nouvelles histones (Inheritance of epigenetic transcriptional memory through read–write replication of a histone modification 2023).
L'action coordonnée de ces protéines permet de reconstituer progressivement le paysage épigénétique caractéristique de chaque région chromosomique (Epigenetic Inheritance: Histone Bookmarks Across Generations 2015).
(Figure : vetopsy.fr modifiée d'après Brickner)
2. La transmission des modifications des histones contribue ainsi, avec le maintien de la méthylation de l'ADN, à la mémoire épigénétique en préservant les états d'expression génique au cours des divisions cellulaires (Functions and mechanisms of epigenetic inheritance in animals 2018).
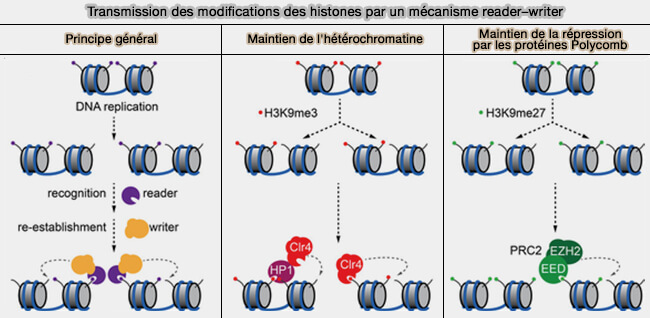
Modèles de mémoire épigénétique
Deux modèles principaux ont été proposés pour expliquer la transmission des états chromatiniens au cours des divisions cellulaires (The molecular principles of gene regulation by Polycomb repressive complexes 2021).
1. Dans le modèle instructif, les marques épigénétiques portées par les nucléosomes parentaux servent directement de modèle au recrutement des protéines readers et des complexes writers.
Les writers rétablissent les mêmes modifications sur les nouvelles histones, assurant ainsi une transmission fidèle des états chromatiniens et une mémoire épigénétique stable.,
2. Dans le modèle adaptatif, les états épigénétiques résultent d'un équilibre dynamique entre les marques de la chromatine, les facteurs de transcription et les réseaux de régulation.
La propagation des marques dépend du contexte cellulaire et de boucles de rétroaction positives ou négatives, permettant de maintenir un état d'expression génique tout en conservant une certaine plasticité.
(Figure : vetopsy.fr d'après Blackledge et Closes)
Ces deux modèles ne sont pas exclusifs.
Les données actuelles indiquent que la mémoire épigénétique résulte généralement de la combinaison
- de mécanismes instructifs, qui assurent la propagation directe des marques épigénétiques,
- de mécanismes adaptatifs, qui stabilisent ou modulent ces états en fonction du contexte biologique.
Bookmarking mitotique
Au cours de la mitose, la chromatine est fortement condensée et la transcription est presque totalement interrompue.
1. Toutefois, certains gènes conservent des repères moléculaires qui facilitent leur réactivation après la division cellulaire (Remembering the past: Mitotic bookmarking in a developing embryo 2018).
Ce mécanisme appelé " bookmarking " (gene bookmarking ou mitotic bookmarking, littéralement " marque-page "), correspond au maintien, sur des régions spécifiques du génome, de facteurs de transcription, de certaines marques d'histones ou d'un état local de la chromatine malgré la condensation des chromosomes.
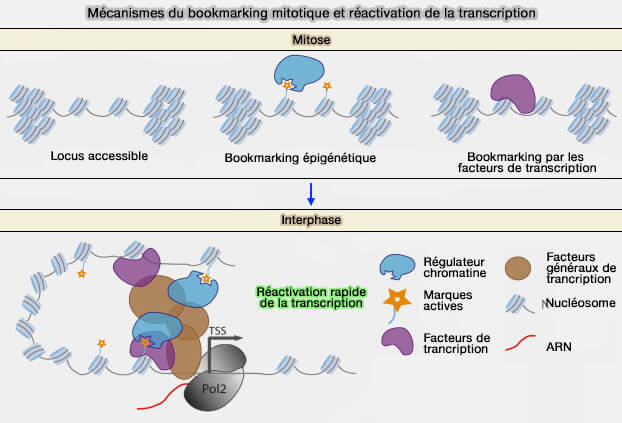
(Figure : vetopsy.fr modifiée d'après Bellec et coll)
2. Après la mitose, ces repères permettent la réactivation rapide des gènes concernés et contribuent à restaurer, dans les cellules filles, le programme d'expression génique caractéristique de chaque type cellulaire.
Le bookmarking mitotique constitue ainsi l'un des mécanismes participant à la mémoire épigénétique.
Reprogrammation épigénétique
Contrairement à la mémoire épigénétique, qui assure la stabilité des programmes d'expression génique, les états épigénétiques peuvent être profondément remodelés au cours :
- du développement embryonnaire,
- de la différenciation cellulaire,
- en réponse à certains stimuli physiologiques ou environnementaux.
Cette capacité d'adaptation, appelée plasticité épigénétique, repose principalement sur la reprogrammation des marques épigénétiques et de l'organisation de la chromatine.
Principes généraux
La reprogrammation épigénétique correspond à l'effacement partiel ou complet des marques épigénétiques préexistantes, suivi de l'établissement de nouvelles marques compatibles avec un autre programme d'expression génique, conduisant progressivement à l'acquisition d'une nouvelle identité cellulaire.
- Selon le contexte biologique, cet effacement n'est toutefois pas absolu.
- Certaines marques épigénétiques ou régions du génome échappent à la reprogrammation, tandis que la majorité des autres marques est réécrite de novo afin de mettre en place un nouvel épigénome adapté à la fonction de la cellule.
La reprogrammation épigénétique s'accompagne d'une profonde réorganisation de la chromatine impliquant :
- la déméthylation et la reméthylation de l'ADN,
- la redistribution des modifications post-traductionnelles des histones,
- le remodelage des nucléosomes,
- la réorganisation de l'organisation tridimensionnelle de la chromatine et des interactions entre les régions régulatrices du génome.
Reprogrammation physiologique
Chez les mammifères, la reprogrammation épigénétique comporte deux grandes vagues successives (Epigenetic Reprogramming in Mice and Humans: From Fertilization to Primordial Germ Cell Development 2023).
1. La première survient après la fécondation.
- Les génomes paternel et maternel subissent une déméthylation massive, accompagnée d'un profond remodelage de la chromatine.
- Cette reprogrammation permet d'effacer une grande partie des marques épigénétiques héritées des gamètes et de rétablir progressivement un état compatible avec la pluripotence des cellules embryonnaires.
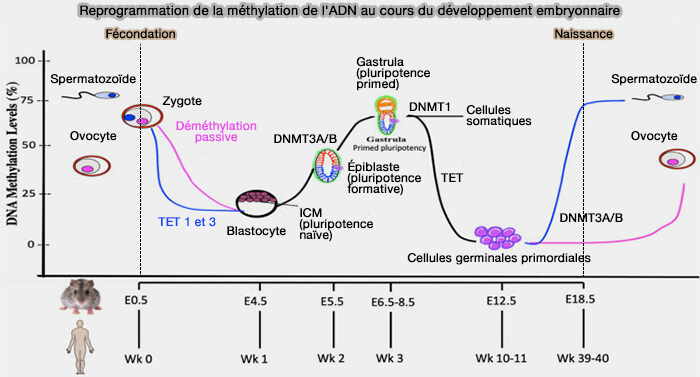
(Figure : vetopsy.fr modifiée d'après Singh et coll)
2. La seconde intervient plus tard, au cours du développement embryonnaire, lors de la formation des cellules germinales primordiales.
Les marques épigénétiques acquises au cours du développement sont de nouveau largement effacées, notamment au niveau de la méthylation de l'ADN, avant qu'un nouvel épigénome, spécifique des spermatozoïdes ou des ovocytes, ne soit progressivement établi au cours de la gamétogenèse.
Reprogrammation expérimentale
La reprogrammation épigénétique peut également être induite expérimentalement (Experimental approaches for the generation of induced pluripotent stem cells 2010).
1. L'expression de facteurs de reprogrammation, notamment les facteurs de Yamanaka (OCT4, SOX2, KLF4 et c-MYC), permet de convertir des cellules différenciées en cellules souches pluripotentes induites (iPS) ou, dans certains cas, de les reprogrammer directement en un autre type cellulaire sans passer par un état pluripotent (Cellular reprogramming and epigenetic rejuvenation 2021).
Cette reprogrammation s'accompagne d'un profond remodelage de la méthylation de l'ADN, des modifications post-traductionnelles des histones, de l'organisation de la chromatine et des profils d'expression génique, conduisant à l'acquisition progressive d'une nouvelle identité cellulaire.
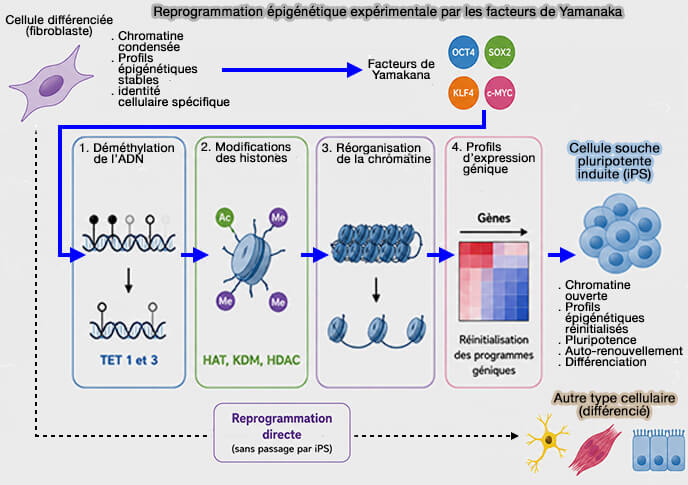
(Figure : vetopsy.fr)
2. Les cellules iPS constituent aujourd'hui un outil majeur de la recherche biomédicale.
Elles permettent de modéliser de nombreuses maladies, d'étudier les mécanismes du développement, de tester de nouvelles molécules thérapeutiques et ouvrent des perspectives importantes en médecine régénérative et en thérapie cellulaire.

La mémoire épigénétique et la plasticité épigénétique constituent deux propriétés complémentaires de l'épigénome, indispensables au maintien de l'identité cellulaire et à l'adaptation des organismes.
La première assure la transmission des programmes d'expression génique au cours des divisions cellulaires, tandis que la seconde permet leur remodelage au cours du développement ou dans des contextes physiologiques et expérimentaux.
BiochimieChimie organiqueBioénergétiqueAcides nucléiquesNucléotidesADNStructureSynthèseOrganisation du génomeRéplicationTranscriptionÉpigénétiqueRéparation de l'ADNARNChromatineNucléosomesHistonesChromosomesProtidesGlucidesLipidesEnzymesCoenzymesVitaminesHormonesComposés inorganiquesTransport membranaireMoteurs moléculairesVoies de signalisation