


Acides nucléiques
ADN : transcription
Activation transcriptionnelle
- Biochimie
- Chimie organique
- Bioénergétique
- Composition de la matière vivante
- Transport membranaire
- Moteurs moléculaires
- Voies de signalisation

L’activation transcriptionnelle regroupe les mécanismes moléculaires permettant le recrutement et l’activation de l’ARN polymérase II grâce aux activateurs transcriptionnels, aux coactivateurs et au remodelage permissif de la chromatine.
Principes généraux de l’activation transcriptionnelle
1. Chez les eucaryotes, l’activation transcriptionnelle dépend généralement de mécanismes coordonnés conduisant à l’établissement d’un environnement chromatinien permissif (euchromatine) associé à une augmentation de l’accessibilité de l’ADN et au recrutement de la machinerie transcriptionnelle, à savoir (Commonly asked questions about transcriptional activation domains 2024) :
- la fixation de facteurs activateurs sur les enhancers ou les éléments proximaux du promoteur,
- le recrutement de coactivateurs transcriptionnels,
- la modification locale de la chromatine,
- la stabilisation des interactions enhancer-promoteur,
- le recrutement du complexe Mediator, de l’ARN polymérase II et du complexe de pré-initiation transcriptionnelle (PIC).
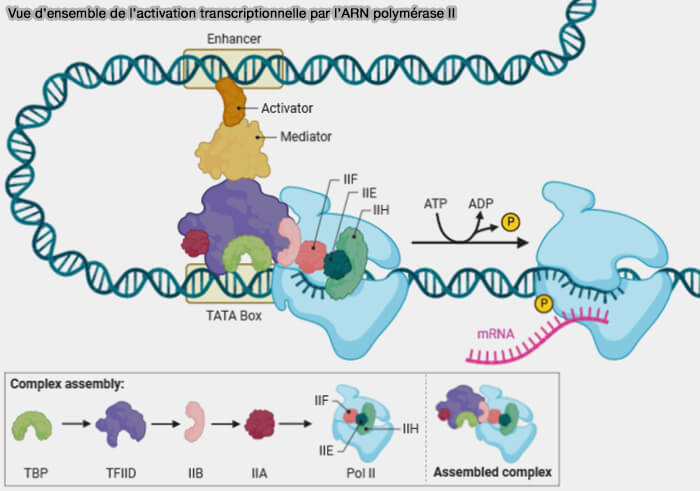
(Figure : vetopsy.fr adaptée d'un template BioRender d'après Shadma Nafis)
2. L’activation transcriptionnelle implique fréquemment :
- l’acétylation des histones par les histones acétyltransférases (HAT),
- l’établissement de marques chromatiniènnes permissives telles que H3K27ac ou H3K4me3,
- le remodelage ATP-dépendant des nucléosomes,
- le déplacement ou l’éjection locale de nucléosomes.
3. Chez les eucaryotes, l’activation transcriptionnelle est également étroitement couplée aux voies de signalisation intracellulaire.
Les activateurs transcriptionnels peuvent agir seuls ou en coopération avec d’autres facteurs de transcription et participent à des réseaux régulateurs complexes dépendant du type cellulaire et des signaux extracellulaires.
De nombreux facteurs de transcription sont régulés par phosphorylation, acétylation, ubiquitination, interactions protéiques, translocation nucléaire induite par des signaux extracellulaires.

Ces mécanismes permettent à la cellule d’adapter rapidement ses programmes transcriptionnels aux variations de l’environnement, aux signaux hormonaux, aux facteurs de croissance, aux stress cellulaires ou aux processus de différenciation.
Acteurs de l'activation transcriptionnelle
Facteurs activateurs
Les facteurs activateurs sont des facteurs de transcription spécifiques capables d’augmenter l’activité transcriptionnelle de gènes cibles par le recrutement de coactivateurs et de la machinerie transcriptionnelle.
Ces protéines reconnaissent des séquences spécifiques d’ADN présentes au niveau des promoteurs, des enhancers ou d’autres éléments cis-régulateurs.
1. La majorité des facteurs activateurs possèdent une organisation modulaire comprenant :
- un domaine de liaison à l’ADN reconnaissant des séquences spécifiques,
- un domaine d’activation transcriptionnelle capable de recruter différents coactivateurs et complexes régulateurs impliqués notamment dans le remodelage de la chromatine, les modifications des histones et l’activation de la machinerie transcriptionnelle.
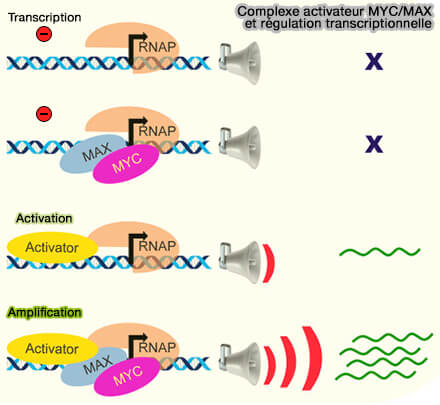
(Figure : vetopsy.fr d'après Jha et coll)
2. Les principaux activateurs transcriptionnels comprennent :
- les complexes Myc/MAX impliqués dans la prolifération et la croissance cellulaire (MYC function and regulation in physiological perspective 2023),
- les complexes OCT4/SOX2 associés au maintien de la pluripotence,
- les complexes AP-1 (Activator Protein 1), constitués de dimères formés par différentes protéines des familles JUN (c-Jun, JunB, JunD) et FOS (c-Fos, FosB, Fra-1, Fra-2), activés notamment par les voies MAPK,
- les facteurs NF-κB impliqués dans les réponses inflammatoires et immunitaires,
- CREB activé par phosphorylation dépendante de PKA,
- HIF-1α stabilisé en condition d’hypoxie.
{kind=link}
3. L’activité des facteurs activateurs dépend fréquemment de mécanismes régulateurs contrôlant :
- leur expression,
- leur localisation nucléaire,
- leur stabilité,
- leurs modifications post-traductionnelles,
- leurs interactions avec d’autres protéines régulatrices.
Remarque : chez les eucaryotes, l’activité de nombreux activateurs transcriptionnels est directement contrôlée par les voies de signalisation intracellulaire ( intégration des voies de signalisation).
intégration des voies de signalisation).
Coactivateurs transcriptionnels
Principes généraux des coactivateurs transcriptionnels
Les coactivateurs transcriptionnels regroupent différentes protéines ou complexes multiprotéiques capables de stimuler la transcription sans reconnaître directement l’ADN de manière spécifique.
Ces facteurs sont recrutés par les activateurs transcriptionnels fixés sur les promoteurs et les enhancers.
1. Les coactivateurs exercent leurs fonctions par plusieurs mécanismes complémentaires impliquant notamment :
- le recrutement de la machinerie transcriptionnelle,
- la modification locale de la chromatine,
- la stabilisation des interactions enhancer-promoteur,
- l’activation de l’initiation et de l’élongation transcriptionnelle.
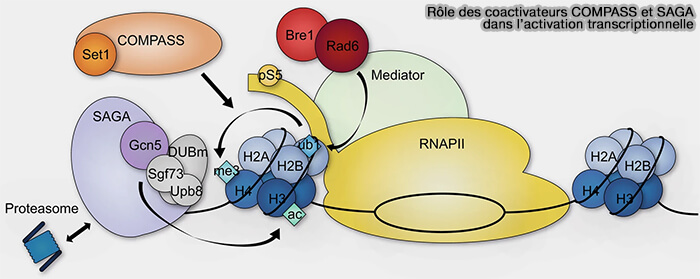
(Figure : vetopsy.fr d'après Cheon et coll)
2. Parmi les principaux coactivateurs figurent notamment :
- les histones acétyltransférases (HAT) telles que CBP et p300 qui favorisent l’acétylation des histones et l’ouverture de la chromatine,
Le complexe SAGA (Spt-Ada-Gcn5 acetyltransferase), un coactivateur transcriptionnel multiprotéique possédant notamment des activités d’acétylation des histones et de déubiquitination impliquées dans l’ouverture de la chromatine et l’activation transcriptionnelle (Dynamic modules of the coactivator SAGA in eukaryotic transcription 2020).
- des complexes possédant des activités de méthylation des histones comme COMPASS impliqué notamment dans les marques H3K4me associées aux régions transcriptionnellement actives,
- les complexes de remodelage chromatinien ATP-dépendants comme SWI/SNF capables de modifier l’organisation des nucléosomes et l’accessibilité de l’ADN,
- le complexe Mediator qui assure l’intégration fonctionnelle entre les facteurs activateurs et la machinerie transcriptionnelle associée à l’ARN polymérase II,
- certains complexes coactivateurs associés aux récepteurs nucléaires comme SRC-1 (The Function of Steroid Receptor Coactivator-1 in Normal Tissues and Cancer 2012).
Cas particulier du complexe Mediator
1. Mediator constitue un complexe coactivateur multiprotéique central de la transcription des gènes transcrits par l’ARN polymérase II (Mediator structure and function in transcription initiation 2023).
- Chez les eucaryotes, ce complexe assure l’intégration fonctionnelle entre les facteurs de transcription spécifiques liés aux enhancers ou aux promoteurs, l’état chromatinien local (euchromatine) et l'assemblage du complexe de pré-initiation transcriptionnelle (PIC).
- Mediator fonctionne comme une plateforme d’interactions dynamique capable d’intégrer simultanément de multiples signaux régulateurs provenant de différentes voies de signalisation et de nombreux facteurs de transcription.
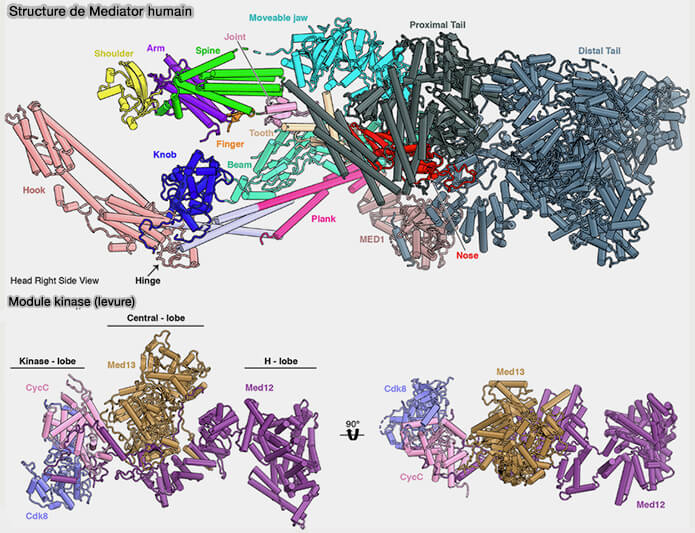
(Figure : vetopsy.fr d'après Rengachari et coll)
2. Le complexe Mediator possède une organisation modulaire comprenant quatre grandes régions composées de nombreux modules et sous-domaines dont la nomenclature varie selon les études structurales, certaines utilisant les sous-unités MED1 à MED30, tandis que d’autres décrivent différentes régions morphologiques comme finger, arm, knob, hook ou plank… (Regulation of the RNA polymerase II pre-initiation complex by its associated coactivators 2024) .
Les grandes régions comprennent :
- un module tête (head) impliqué dans les interactions avec l’ARN polymérase II, notamment avec le domaine CTD (C-terminal domain) de la sous-unité RPB1, ainsi qu’avec le complexe de pré-initiation transcriptionnelle (PIC),
- un module central (middle) assurant une partie de l’organisation structurale du complexe et participant à la transmission des signaux régulateurs entre les activateurs transcriptionnels et l’ARN polymérase II,
- un module queue (tail) interagissant avec de nombreux activateurs transcriptionnels liés aux enhancers et contribuant au recrutement de Mediator ainsi qu’à l’organisation des interactions entre enhancers et promoteurs,
- un module kinase (CKM) dissociable contenant notamment CDK8, la cycline C et MED12 et MED13, capable de moduler positivement ou négativement l’activité transcriptionnelle par phosphorylation de différentes protéines impliquées dans la régulation de la transcription, notamment au niveau de l’initiation, de la pause proximale et de l’élongation transcriptionnelle.
Remarque : le module kinase de Mediator diffère du complexe CAK (CDK-activating kinase) associé à TFIIH et peut coopérer fonctionnellement avec celui-ci dans la régulation de la transcription par l’ARN polymérase II.
Cependant, contrairement au CAK principalement impliqué dans l’activation transcriptionnelle, le module kinase contenant CDK8 peut exercer des effets activateurs ou inhibiteurs selon le contexte transcriptionnel et les facteurs régulateurs associés.

(Figure : vetopsy.fr d'après Rengachari et coll)
Mécanismes d’activation transcriptionnelle
Remodelage activateur de la chromatine et modifications des histones
L’activation transcriptionnelle nécessite fréquemment l’établissement local d’un environnement chromatinien permissif (euchromatine) favorisant l’accessibilité des promoteurs, des enhancers et d'autres éléments cis-régulateurs aux facteurs de transcription et à la machinerie transcriptionnelle.
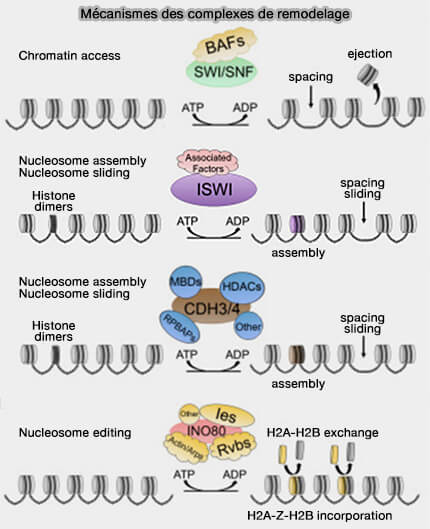
(Figure : vetopsy.fr d'après Magaña-Acosta et coll)
Chez les eucaryotes, cette ouverture de la chromatine dépend notamment de complexes de remodelage ATP-dépendants et de modifications post-traductionnelles des histones qui agissent de manière coordonnée.
1. Les complexes de remodelage chromatinien ATP-dépendants utilisent l’énergie issue de l’hydrolyse de l’ATP pour modifier l’organisation des nucléosomes et peuvent :
- déplacer les nucléosomes le long de l’ADN,
- modifier leur espacement,
- favoriser leur éjection locale,
- augmenter l’accessibilité de régions régulatrices spécifiques.
Les complexes SWI/SNF (BAF chez les mammifères), ISWI, CHD, INO80. sont étudiés dans un chapitre spécifique.
2. L’activation transcriptionnelle est également associée à différentes modifications activatrices des histones qui favorisent généralement l’ouverture de la chromatine et le recrutement de protéines régulatrices ( code des histones).
a. Les modifications des histones et les mécanismes de remodelage chromatinien sont étroitement couplés, notamment par :
- l’acétylation des histones par les histones acétyltransférases (HAT) telles que CBP/p300 ou GCN5 sur différentes lysines, modification qui neutralise partiellement les charges positives des histones, diminue leurs interactions avec l’ADN négativement chargé et favorise le recrutement de protéines possédant des bromodomaines capables de reconnaître ces lysines acétylées et de stabiliser les états chromatiniens actifs,
- certaines méthylations de lysines comme H3K4me3 fortement enrichie au niveau des promoteurs actifs, H3K4me1 fréquemment associée aux enhancers et H3K36me3 enrichie dans les régions transcrites lors de l’élongation transcriptionnelle,
- certaines phosphorylations comme H3S10ph ou H3S28ph et ubiquitinations comme H2Bub1 associées aux états chromatiniens actifs.
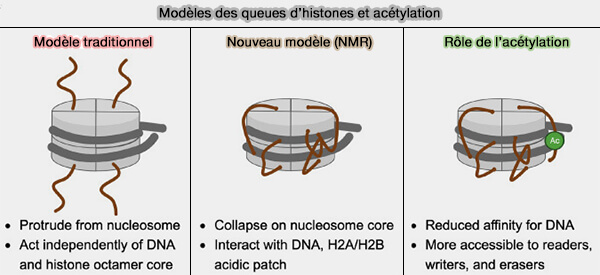
(Figure : vetopsy.fr d'après Weinzapfel et coll)
b. L’établissement, la reconnaissance et l’élimination de ces modifications reposent respectivement sur des enzymes writers, des protéines readers et des enzymes erasers ( principe combinatoire des modifications d’histones).
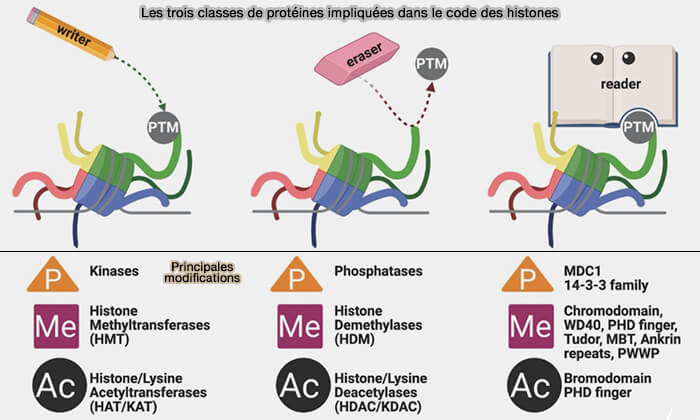
(Figure : vetopsy.fr d'après Malone et coll)
3. Ces mécanismes facilitent :
- la fixation des facteurs de transcription sur les régions régulatrices accessibles,
- le recrutement des coactivateurs transcriptionnels, du complexe Mediator et de l’ARN polymérase II,
- la stabilisation des interactions enhancer-promoteur,
- l’activation de l’initiation transcriptionnelle et le passage vers une élongation productive.
Recrutement et activation de l’ARN polymérase II
L’activation transcriptionnelle conduit finalement au recrutement et à l’activation de l’ARN polymérase II au niveau des promoteurs des gènes transcrits.
1. Chez les eucaryotes, ce processus dépend de l’assemblage coordonné de facteurs généraux de transcription, de coactivateurs transcriptionnels et du complexe Mediator qui stabilisent le recrutement de la machinerie transcriptionnelle au niveau des promoteurs et des enhancers.
Les mécanismes détaillés d’assemblage du complexe de pré-initiation transcriptionnelle (PIC) et d’initiation transcriptionnelle sont étudiés dans la page consacrée au mécanisme de la transcription.
2. L’activation transcriptionnelle dépend également du contrôle de la transition entre initiation et élongation productive (pause proximale de l’ARN polymérase II).
- La pause proximale est induite par les facteurs NELF et DSIF.
- La levée de cette pause dépend principalement du complexe P-TEFb contenant CDK9 qui phosphoryle le CTD de l’ARN polymérase II ainsi que NELF et DSIF, favorisant la dissociation de NELF, la conversion de DSIF en facteur positif d’élongation et l’entrée en élongation productive.
3. Chez les eucaryotes, de nombreux gènes régulés possèdent une ARN polymérase II déjà recrutée au niveau du promoteur mais temporairement bloquée peu après le début de la transcription par des mécanismes de pause proximale.
La levée rapide de cette pause permet alors une entrée rapide en élongation productive en réponse aux signaux cellulaires ( chapitre suivant).
Intégration des voies de signalisation
Chez les eucaryotes, les voies de signalisation intracellulaire modulent directement l’activité transcriptionnelle par l’activation, la stabilisation ou la translocation nucléaire de différents facteurs de transcription et coactivateurs.
1. La régulation transcriptionnelle permet à la cellule d’adapter rapidement ses programmes d’expression génique aux variations de l’environnement.
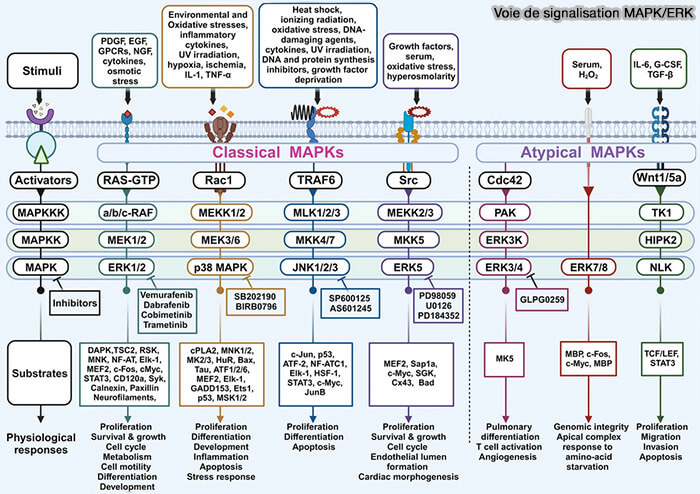
a. Les signaux extracellulaires tels que les facteurs de croissance, cytokines, hormones, stress cellulaires ou variations d’oxygénation activent différentes cascades de signalisation capables de modifier l’activité de nombreux facteurs de transcription et coactivateurs.
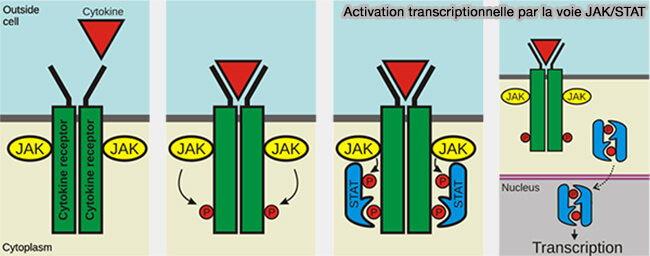
(Figure : vetopsy.fr d'après Peter Znamenkiy)
{kind=link}
b. Par exemple :
- CREB est activé par phosphorylation dépendante de PKA,
- STAT est activé par les voies JAK/STAT,
- NF-κB est libéré de IκB puis transloqué dans le noyau,
- SMAD est activé par la signalisation TGF-β,
- HIF-1α est stabilisé en condition d’hypoxie.
2. Plusieurs voies de signalisation peuvent converger vers les mêmes enhancers ou promoteurs et moduler de manière coordonnée l’activité transcriptionnelle.
Répression de la transcription
BiochimieChimie organiqueBioénergétiqueAcides nucléiquesNucléotidesADNStructureSynthèseOrganisation du génomeRéplicationTranscriptionÉpigénétiqueRéparation de l'ADNARNChromatineNucléosomesHistonesChromosomesProtidesGlucidesLipidesEnzymesCoenzymesVitaminesHormonesComposés inorganiquesTransport membranaireMoteurs moléculairesVoies de signalisation