Elle internalise un grand volume de matériel, dans des vésicules de tailles supérieures à 200 nm, appelées macropinosomes.
Replis membranaires dorsaux circulaires
(CDR : Circular Dorsal Ruffle)
(Figure : vetopsy.fr d'après Lin et coll)
Ce processus, à base d'actine, hautement conservé par l'évolution, est un moyen pour les cellules du système immunitaire inné d'inspecter les antigènes/les molécules microbiennes associées (The origins and evolution of macropinocytosis 2018).
Ce type d’endocytose repose sur la génération de replis membranaires (ruffles) qui vont se refermer en fusionnant pour former des macropinosomes.
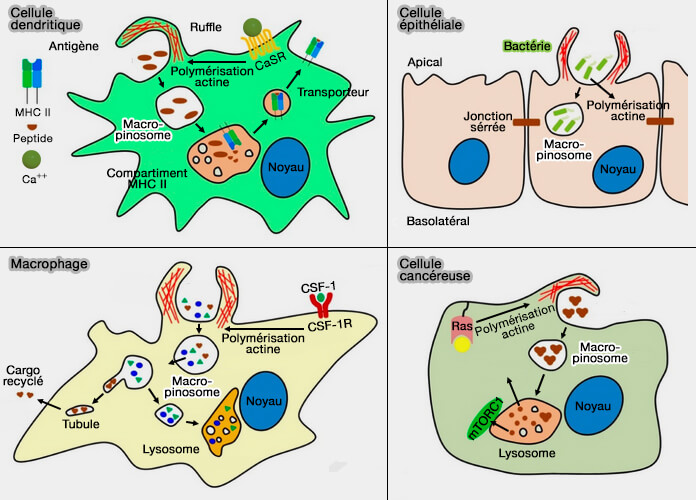
2. La macropinocytose intervient dans plusieurs processus dans la plupart des types cellulaires (SnapShot : Macopinocytosis 2017) :
la surveillance immunitaire, i.e. pour reconnaître les antigènes ou les molécules associées aux agents infectieux,
lors d'une signalisation élevée et soutenue induite par des facteurs de croissance, par la chimiokine, par des récepteurs Toll-like (TLR) ou par des ligands d'intégrines,
par des restes cellulaires apoptotiques et certains virus.
Macropinocytose dans différentes cellules
(Figure : vetopsy.fr d'après Lin et coll)
Remarque : dans les synapses, l'ADBE (endocytose de masse dépendante de l'activité) est le mode endocytaire dominant et partage les caractéristiques de la macropinocytose, vésicules induisant des invaginations membranaires, engloutissement non spécifique de toutes les molécules situées sur la plaque de la membrane plasmique utilisées pour former les grands " bulk endosomes ".
1. La différence avec la phagocytose est que la phagocytose :
engloutit des contenus solides,
est une réponse immunitaire,
est localisée dans la plupart des cellules immunitaires.
1. Dans le cas de l'engagement des récepteurs opsoniques reconnaissant des anticorps, des compléments ou des lectines autour de la surface des pathogènes opsonisés ou des cellules apoptotiques, la macropinocytose rappelle la phagocytose.
a. Toutefois, elle n'entre pas en interaction directe avec les cargos, mais peut engloutir :
le patch membranaire contenant ces cargos,
un grand volume de fluide extracellulaire.
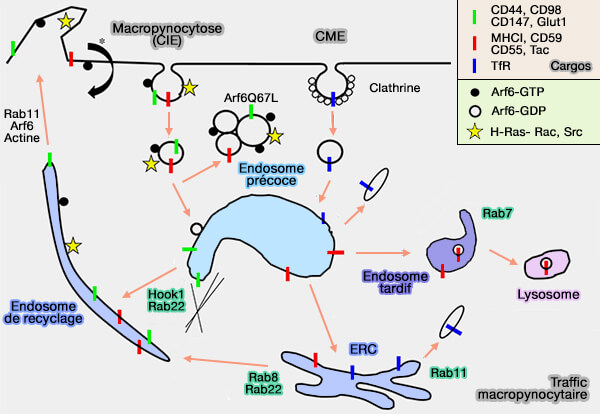
b. Il n'y a pas de tri des mécanismes à la surface de la cellule, et le destin des récepteurs intériorisés, i.e. recyclage à la surface ou dégradation dans les lysosomes, est décidé par la maturation endosomale.
La macropinocytose est activée par des signaux spécifiques, mais n'est pas spécifique au cargo et de nombreuses protéines transmembranaires sont internalisées en sus.
Mécanisme de formation
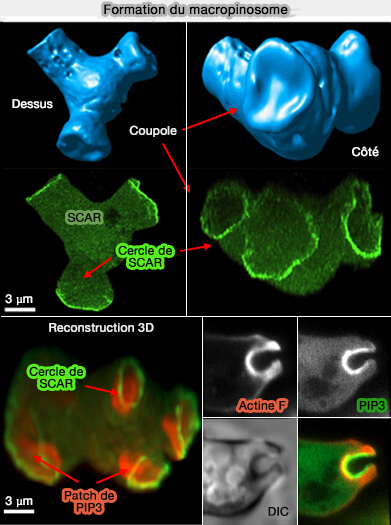
1. Dans la macropinocytose, des replis ou protrusions membranaires (ruffles) en forme de coupole, souvent appelés replis membranaires dorsaux circulaires (CDR : Circular Dorsal Ruffle), forment des protubérances à la surface des cellules.
Bien que de nombreux ruffles disparaissent peu de temps après leur formation, certains ruffles en forme de feuille se transforment en ruffles circulaires.
Les bords avant de ces ruffles grandissent par polymérisation de l'actine et doivent être poussés vers l'extérieur pour enfermer le contenu extracellulaire, alors que leur base doit être maintenue statique.
Les cupules résultantes peuvent mesurer plusieurs microns de diamètre et finir par se fermer par constriction de leur bord, la fusion membranaire produisant une vésicule endocytaire.
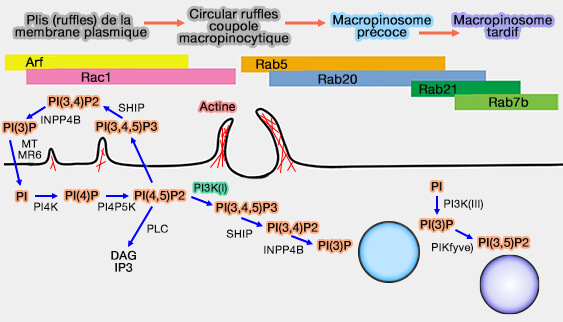
Formation de macropinosome
(Figure : vetopsy.fr d'après Veltmann et coll)
c. L'activation de Rac-GTP et de Cdc42-GTP stimule également PAK1 (p21-activated kinase 1), ce qui soutient la réorganisation de l'actine et la formation des ruffles.
3. La formation de la coupole d'actine est activée par un patch Ras/PI(3,4,5)P3 central, invariablement, associé à un anneau de WAVE ou à son homologue SCAR, actif à son bord, qui a un rôle essentiel dans la nucléation de l'actine (A plasma membrane template for macropinocytic cups 2016 avec de nombreuses vidéos explicatives).
L'interaction électrostatique entre PI(3,4,5)P3 et les motifs polybasiques, i.e. domaine B, des facteurs de nucléation de l'actine est essentielle au bourgeonnement membranaire.
On peut ainsi bloquer la macropinocytose en empêchant les échanges NA+/H+, conduisant à l'augmentation de la concentration cytosolique de H+ et à la neutralisation des charges négatives de la couche interne membranaire.
Remarque : conformément au rôle central de PI(3,4,5)P3, la régulation s'effectue par des phosphatases comme PTEN, qui reforme le PI(4,5)P2, et SHIP2, qui le transforme en PI(3,4)P2.
5. Les étapes finales de la formation des macropinosomes, i.e. fermeture de la vésicule,sont mal connues.
La désactivation qui suit l'activation de Rac1 serait cruciale pour l'achèvement de la formation des macropinosomes et leur fermeture.
CTBP1/BARS subit une translocation vers la coupe macropinocytaire et sa membrane environnante lors de la stimulation par EGF et a été impliqué dans sa scission.
Certains rapports impliquent la dynamine, mais comme elle possède de nombreux effets en dehors de la macropinocytose, son rôle doit être interprété avec prudence (Dynasore - not just a dynamin inhibitor 2015).






 mécanisme de l'ADBE).
mécanisme de l'ADBE).