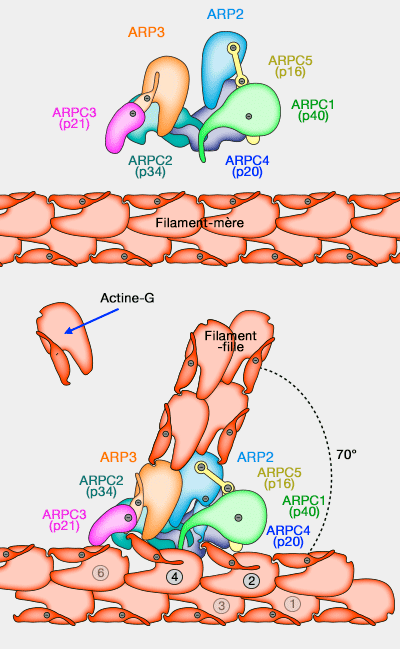
Trafic vésiculaire
Endocytose clathrine-dépendante (CME)
4. Approfondissement du puits recouvert de clathrine (CCP)
4b. Polymérisation de l'actine et dynamine
L'approfondissement du puit recouverts de clathrine (CCP) et le rétrécissement de son cou, i.e. transition Plat ➞ Λ ➞ Ω, forment une poche profondément invaginée, grâce à la polymérisation de l'actine, mais aussi à la dynamine.
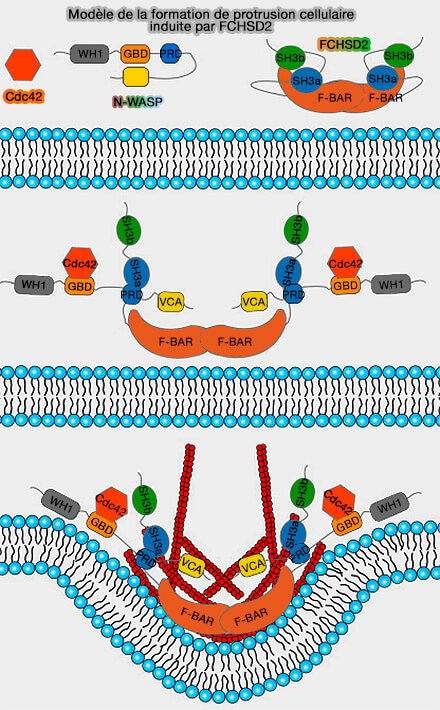
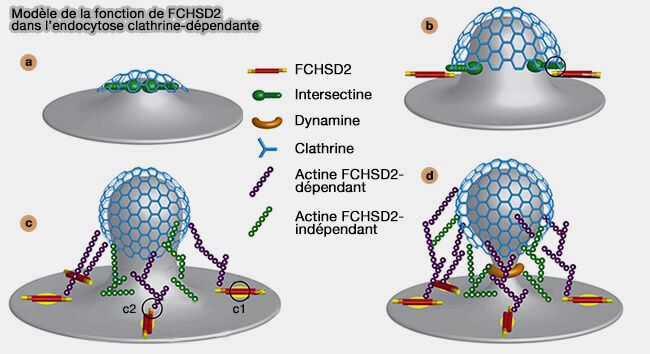
b. FCHSD2 est recrutée par l'intersectine par son interaction entre les domaines SH3, et la formation de protrusion cellulaire induite par FCHSD2 nécessite sa capacité de liaison à la membrane.
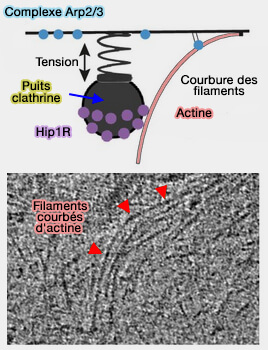
1. Une fraction significative de microfilaments d'actine se plie sous la force générée par la polymérisation et contribue au succès de l'internalisation endocytaire d'au moins deux manières.
Ces filaments courbés stockent de l'énergie élastique pour la production de force ultérieure, tout comme la perche d'un sauteur.
Courbure des filaments d'actine
(Figure : vetopsy.fr d'après
Akamatsu et coll)
2. Ces filaments courbés contribuent à l'internalisation en libérant l'énergie élastique stockée lorsqu'ils se redressent sous des fluctuations thermiques, conformément au mécanisme élastique à cliquet brownien pour la production de force médiée par l'actine (Actin Mechanics and Fragmentation 2015).
Une partie de l'énergie élastique des filaments courbés est libérée à mesure que l'internalisation augmente, suggérant une compression réversible du réseau pour stocker l'énergie élastique.
Dans l'endocytose, la distribution des Hip1R autour du puits dirige davantage de filaments vers la base du puits, ce qui nucléé plus de filaments de manière autocatalytique et augmente la flexion des filaments, favorisant ainsi une plus grande internalisation.
Les réseaux de microfilaments d'actine et les ABP (Actin Binding Protein) provoquent, par leur croissance, l'invagination des vésicules d'endocytose, leur intériorisation et leur transport dans la cellule.
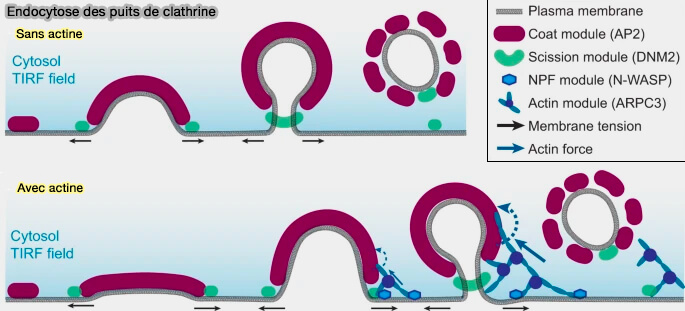
1. Lorsque les conditions locales sont favorables à la déformation de la membrane par les protéines de l'enveloppe, la membrane peut subir une transition Plat ➞ Λ ➞ Ω dans un temps relativement court sans assemblage d'actine.
Lorsque le manteau devient suffisamment grand pour former un bourgeon en forme de Ω, la dynamine peut être recrutée pour effectuer la scission, et il y a peu de délai entre l'expansion du manteau et la scission.
Deux endocytoses des puits de clathrine dans la CME
(Figure : vetopsy.fr d'après Jin et coll)
2. Lorsque les conditions locales ne sont pas favorables, probablement sous une tension membranaire élevée ou d'autres obstacles, l'interaction protéine d'enveloppe-membrane ne génère pas une force suffisante pour courber la membrane.
Le recrutement asymétrique de N-WASP active la nucléation de l'actine principalement sur un côté de l'enveloppe de clathrine, générant une force asymétrique qui attire la membrane dans la cellule comme un décapsuleur.
c. Les événements CME associés à l'assemblage d'actine ont des durées de vie plus longues, probablement en raison du recrutement adaptatif de régulateurs d'actine suivi d'un assemblage de réseaux d'actine et d'un remodelage de la membrane.
Ces résultats établissent l'hétérogénéité des sites pour la dépendance et l'implication de l'actine dans la CME.
Et la dynamine
La dynamine et l'actine peuvent exercer leurs forces de traction et de constriction pour générer des puits enrobés de clathrine.
La dynamine pourrait être prérecrutée dans des zones endocytaires par interaction avec la syndapine 1 ( dynamine et UFE).
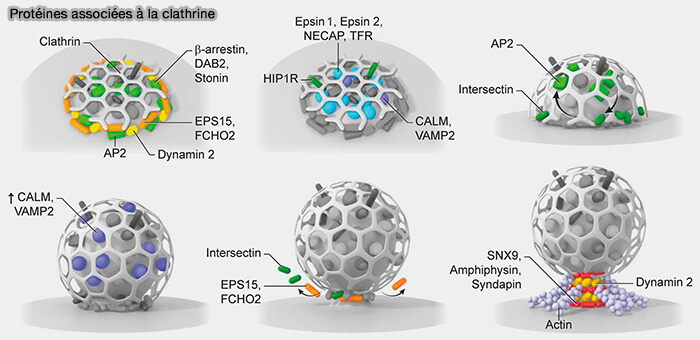
Protéines associées à la clathrine dans la CME
(Figure : vetopsy.fr d'après Sochaki et coll)
La traction et la constriction dépendantes de l'actine et de la dynamine sous-tendent la transformation du profil Plat ➞ Ο, indépendamment de l'implication de protéines de revêtement comme la clathrine.




 approfondissement de CCP et dynamine).
approfondissement de CCP et dynamine).