Complexe APC/C
Régulation extrinsèque
2. Point de contrôle du fuseau mitotique (SAC)
2.1 Prométaphase : activation du SAC
b. Structure de Mad1, Mad2 et leur interaction
Le complexe Mad1-Mad2 constitue le cœur catalytique du point de contrôle du fuseau, convertissant Mad2 en sa forme active et amorçant la formation du signal inhibiteur du SAC.
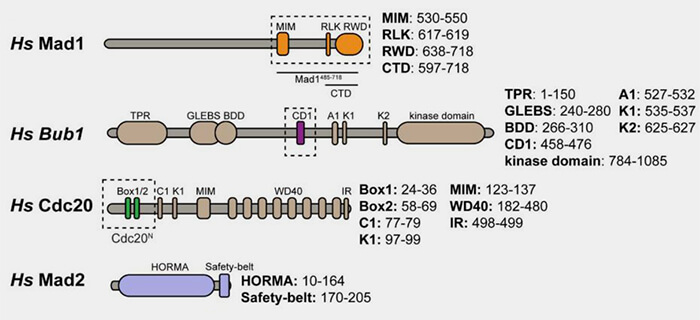
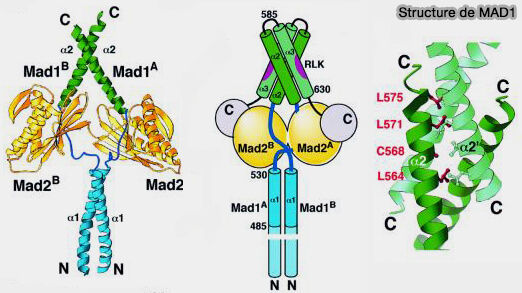
Structure de MAD1, MAD2, BUB1 et Cdc20
(Figure : vetopsy.fr d'après Fischer et coll)
Structure de MAD1 et MAD2
Sur la plateforme formée par BUB1 au niveau de KNL1, le complexe MAD1-MAD2 constitue le cœur du mécanisme de conversion du signal du SAC, transformant l’absence d’attachement en signal inhibiteur.
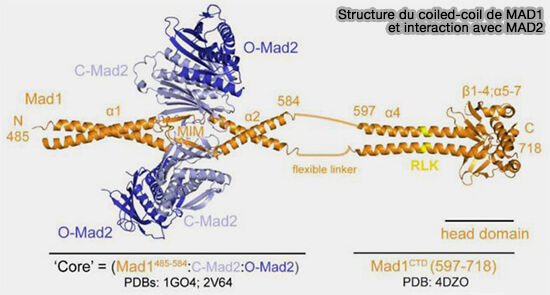
Ce segment est suivi d'un lien flexible (résidus 585-597), après quoi la bobine enroulée reprend.
2. Ce domaine forme un échafaudage rigide qui maintient les deux extrémités C-terminales proches l’une de l’autre, permettant la fixation d'une molécule de Mad2 à chaque extrémité.
Structure du coiles-coil de MAD1 et interactions avec O-Mad2
(Figure : vetopsy.fr d'après Fischer et coll)
Domaine C-terminal
Le domaine C-terminal (597-718) regroupe l’extrémité des hélices α dans laquelle on peut différencier deux modules structuraux distincts.
1. Un motif RLK (617-619), i.e. arginine-leucine-lysine, reconnaît Bub1 phosphorylé pour assurer le recrutement stable du complexe Mad1-Mad2 au kinétochore.
Ce domaine adopte une architecture en sandwich β-α-β, composée de quatre feuillets β (β1-β4) disposés autour de deux hélices α, formant un noyau hydrophobe compact.
a. Bien qu’il ne possède pas d’activité catalytique, il joue un rôle stabilisateur et adaptateur.
Il oriente la région CD1 phosphorylée vers le complexe Mad1-Mad2.
Il contribue à la topologie correcte du module Bub1-Bub3-Mad1 au kinétochore.
b. Ainsi, la tête RWD constitue un pivot structurel essentiel, assurant la connexion mécanique et fonctionnelle entre la portion coiled-coil de Bub1 et sa région d’interaction avec Mad1.
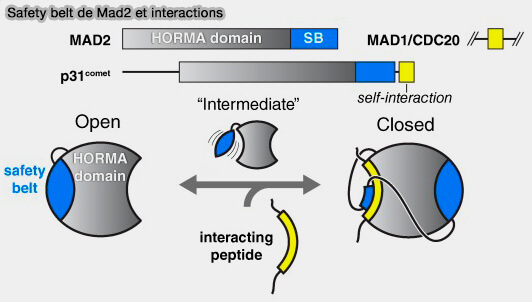
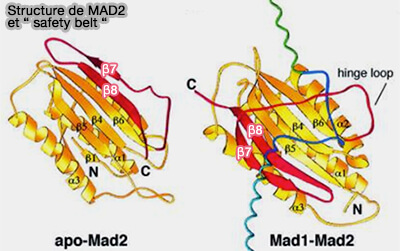
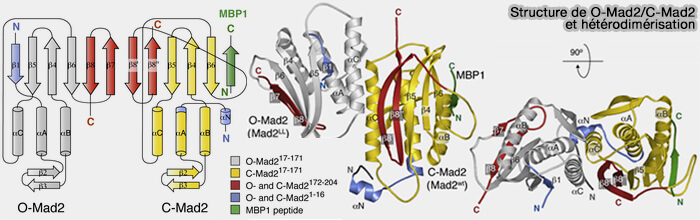
a. O-Mad2 (ouverte) est la forme inactive cytosolique.
La chaîne β5-β6 est non repliée et la région C-terminale (feuillet β7 et boucle adjacente) est libre et ne recouvre pas le ligand.
La poche hydrophobe de Mad2 est accessible, prête à accueillir un peptide cible (comme le motif MBM de Mad1 ou celui de Cdc20).
b C-Mad2 (fermé) se forme lorsque Mad2 reconnaît un peptide partenaire, le motif MIM/MBM de Mad1 ou Cdc20.
Le segment C-terminal bascule autour du ligand et se replie sur lui comme une ceinture de sécurité, d’où le nom de " safety-belt ".
Ce repliement fait passer la β7 et la boucle adjacente au-dessus du ligand, piégeant littéralement le peptide dans la poche.
La fermeture crée une barrière stérique rendant la dissociation du ligand extrêmement lente : c’est une interaction quasi irréversible à l’échelle mitotique.
Safety belt de Mad2 et interactions
(Figure : vetopsy.fr d'après Rosenberg et Corbett)
2. Mad2 entre en interaction avec plusieurs partenaires :
avec p31comet, pour la conversion inverse C-Mad2/O-Mad2, essentielle à la désactivation du checkpoint.
Interaction Mad1-Mad2
1. La rencontre entre MAD1 et O-MAD2 s'effectue au pore nucléaire.
a. Durant l’interphase, Mad1 et Mad2 ne sont pas encore localisés aux kinétochores.
Structure de Mad2
(Figure : vetopsy.fr d'après Zhang et coll)
Mad1 est associée de manière constitutive à certaines protéines des pores nucléaires comme Tpr (Translocated Promoter Region) et Nup153, formant un complexe périnucléaire stable en forme de panier.
Mad2, sous sa forme ouverte (O-Mad2), circule librement dans le cytoplasme et le nucléoplasme.
Mad1 se lie à la région N-terminale de TPR, tandis que Mad2 se lie à sa région C-terminale.
La phosphorylation de TPR au niveau du résidu S2059 par CDK1 est cruciale pour l’interaction TPR-Mad1.
La perturbation de la phosphorylation de S2059 abolit cette interaction, compromettant la localisation de Mad1 et de Mad2, induisant ainsi des anomalies du cycle cellulaire.
Le complexe Mad1-Mad2 observé aux pores est structurellement identique à celui retrouvé plus tard aux kinétochores non attachés.
2. La région C-terminale de Mad1 contient un motif MIM/MBD, séquence courte (~20 aa) contenant un peptide hydrophobe (ΦxxΦxΦ) qui s’insère dans la poche β-sandwich de O-Mad2 qui subit un changement conformationnel majeur.
O-Mad2 referme sa " safety-belt ", ce qui déclenche sa fermeture et sa transformation en C-Mad2.
Remarque : la fermeture de Mad2 est encore sujette à caution.
Ce processus serait une conversion allostérique spontanée, déclenchée uniquement par la liaison de Mad1.
3. Une fois Mad2 refermée, elle reste solidement associée à Mad1.
Mad1 s’homodimérise par sa longue coiled-coil ce qui permet la fixation de deux molécules de C-Mad2, une à chaque extrémité.
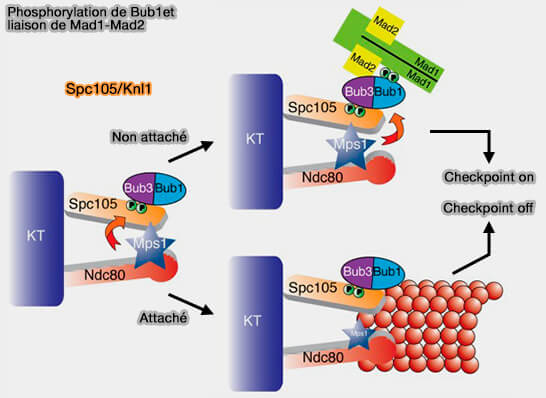
Ce complexe préformé est stable tout au long de la prophase et sera relocalisé aux kinétochores non attachés après la rupture de l’enveloppe nucléaire, grâce à l’action de MPS1 qui phosphoryle Bub1.
Cette structure est la base du modèle catalytique du SAC : Mad1-C-Mad2 agit comme une enzyme allostérique et est à la base de son mécanisme d’amplification.
Phosphorylation de Bub1 et liaison de Mad1-Mad2
(Figure : vetopsy.fr d'après London et Biggins)




 implications of a ‘safety belt’ binding mechanism for the spindle checkpoint 2002).
implications of a ‘safety belt’ binding mechanism for the spindle checkpoint 2002).