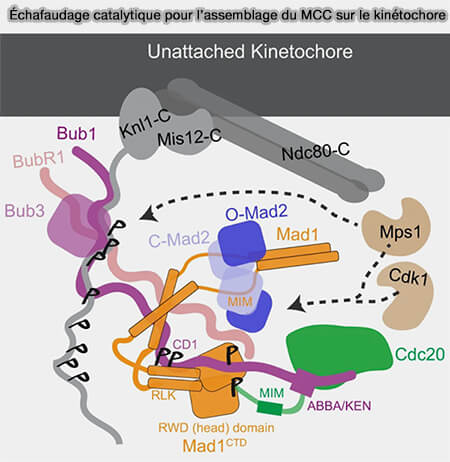
a. La couche interne constitue l’interface avec le centromère et repose sur la chromatine spécialisée.
Elle s’organise autour du nucléosome CENP-A et du réseau CCAN, qui assurent l’ancrage constitutif du kinétochore et la transmission stable de l’identité centromérique au cours des cycles cellulaires.
b. La couche externe correspond au cœur mitotique du kinétochore et établit le lien direct avec les microtubules.
Dominée par le réseau KMN, elle permet l’attachement dynamique aux microtubules, la conversion des variations de longueur en forces de traction et la génération de tension lors de la biorientation des chromosomes.
c. La corona fibreuse forme une structure périphérique transitoire spécialisée dans la capture et la signalisation.
Elle augmente la surface d’interaction avec les microtubules en début de mitose et sert de plateforme pour le recrutement des composants impliqués dans le point de contrôle du fuseau (SAC).
Le kinétochore agit comme un centre d’intégration et traduit l’état des attachements et de la tension en signaux régulant l’activation de l’APC/C et la transition métaphase-anaphase.
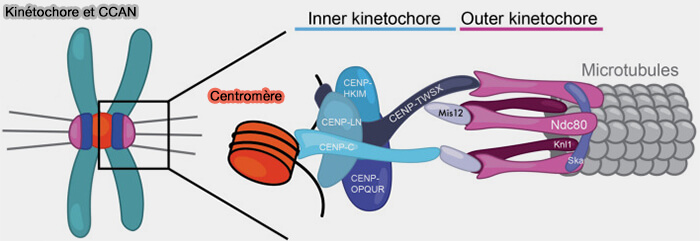
Couche interne : interface avec le centromère
La couche interne du kinétochore constitue la zone de contact stable avec le centromère et fournit la plateforme structurale nécessaire à l’assemblage de la couche externe.
a. Ce nucléosome est formé par CENP-A, qui remplace l'histone H3 canonique et définit l’identité centromérique, ainsi que par les histones H4, H2A et H2B.
Les propriétés structurales de CENP-A confèrent au nucléosome centromérique une stabilité différente de celle des nucléosomes canoniques ( différences structurales avec H3).
Cette organisation particulière induit une plus grande plasticité de l’ADN enroulé, favorisant des conformations compatibles avec l’assemblage du réseau CCAN et l’ancrage du kinétochore.
b. Cette couche intègre le réseau CCAN (Constitutive Centromere-Associated Network) qui assure la continuité structurale entre la couche interne et la couche externe du kinétochore par deux voies principales.
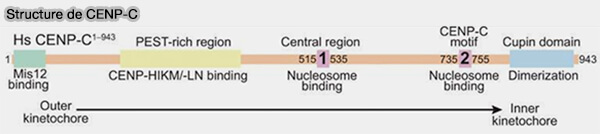
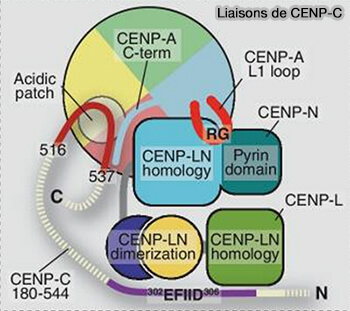
1. CENP-C, qui sert de patron/modèle pour l'assemblage de kinétochore par une succession N- à C-terminale de motifs de liaison alignés le long de l'axe extérieur à l'axe intérieur, entre en interaction avec :
CENP-A par son extrémité C-terminale, assurant l’ancrage du kinétochore à la chromatine centromérique.
le complexe MIS12 (MIS12C) du réseau KMN du kinétochore externe, par son extrémité N-terminale, établissant une connexion mécanique directe entre le centromère et les microtubules du fuseau.
Structure de CENP-C
(Figure : vetopsy.fr d'après Walstein et coll)
L’extrémité N-terminale de CENP-T, qui ne présente pas de domaine de type histone-fold, porte des motifs de liaison directs au complexe Ndc80 ainsi que des sites de phosphorylation dépendants de CDK1, et selon le contexte, de Plk1.
Ces phosphorylations modulent l’affinité de CENP-T pour le complexe Ndc80 et renforcent le recrutement du réseau KMN, de manière indépendante de la voie CENP-C/Mis12, conférant à cette interface un caractère directement régulable au cours de la mitose.
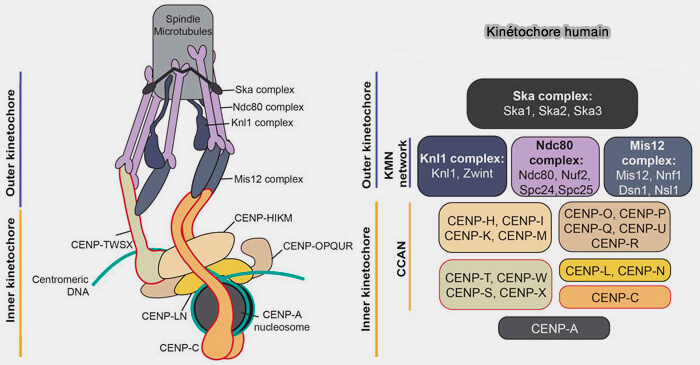
La couche externe du kinétochore établit le lien mécanique et signalétique avec le fuseau mitotique en assurant :
l’attachement des microtubules,
la transmission des forces de traction générées par leur polymérisation et dépolymérisation,
l’intégration des signaux contrôlant la progression de la mitose.
Sur le plan architectural, cette couche s’organise autour de deux modules fonctionnels distincts.
Le réseau KMN (KNL1-Mis12-Ndc80), constitue l’interface structurale stable reliant la chromatine centromérique, via le CCAN, aux microtubules du fuseau,
La corona fibreuse, structure périphérique transitoire, spécialisée dans la capture initiale des microtubules et l’amplification de la signalisation du point de contrôle du fuseau.
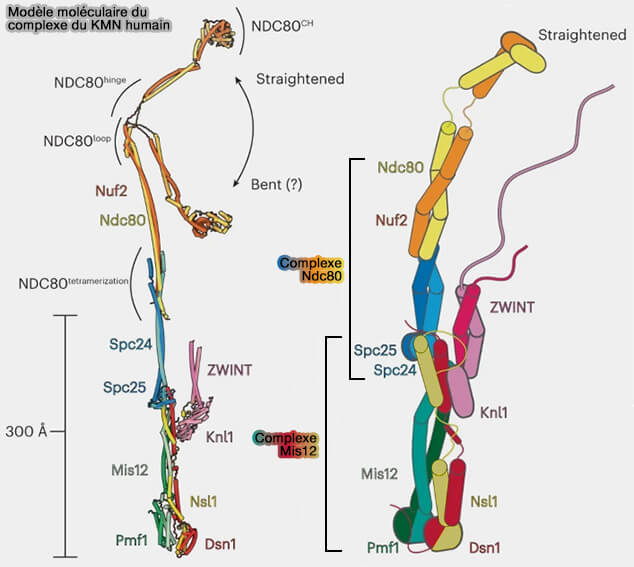
Modèle moléculaire du résaeu KMN humain
(Figure : vetopsy.fr d'après Yatskevich et coll)
Complexe Mis12 (Mis12C)
Le complexe Mis12 (Mis12C), composé de DSN1, MIS12, NSL1 et PMF1, constitue le nœud d’assemblage central du réseau KMN au sein de la couche externe du kinétochore, à l’interface entre le réseau CCAN centromérique et les composants du kinétochore externe impliqués dans l’attachement aux microtubules.
Il assure la transmission structurale de l’ancrage fourni par CENP-C vers les modules d’attachement et de signalisation portés par Ndc80 et KNL1.
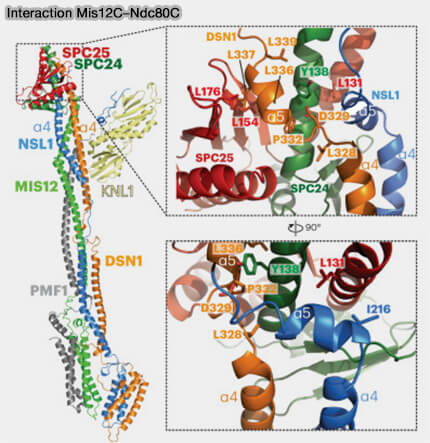
Interaction Mis12C et NDC80C
(Figure : vetopsy.fr d'après Polley et coll)
Sur le plan architectural, Mis12C adopte une organisation allongée et rigide, adaptée à une fonction d’échafaudage.
1. MIS12, NSL1 et PMF1 constituent l’armature structurale du complexe Mis12.
Ils stabilisent l’assemblage du réseau KMN et assurent le positionnement correct de Ndc80 et KNL1 par rapport à la plateforme centromérique.
2. DSN1 occupe une position centrale dans l’interface entre le complexe Mis12 et les différentes plateformes du kinétochore externe.
Son extrémité N-terminale contient des sites de phosphorylation dépendants des kinases mitotiques, notamment Aurora B, qui modulent l’affinité de Mis12C pour CENP-C et conditionnent la stabilité de l’ancrage du kinétochore externe.
Cette régulation permet d’ajuster dynamiquement l’assemblage du KMN en fonction de l’état d’attachement et de tension.
3. Mis12C établit une interface structurale avec le complexe Ndc80, en particulier avec le module basal SPC24-SPC25, assurant l’ancrage du complexe Ndc80 au réseau KMN et le positionnement correct de l’interface d’attachement end-on aux microtubules.
Cette connexion assure le positionnement correct de Ndc80 par rapport à la plateforme centromérique et permet la transmission des forces de traction générées par la dynamique des microtubules.
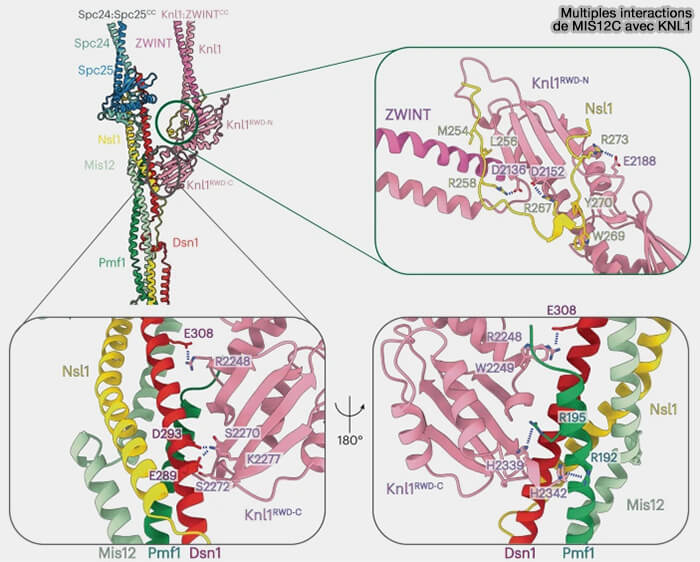
4. Mis12C interagit également par de multiples interactions avec KNL1, via une organisation structurale continue du réseau KMN.
Cette interface permet d’aligner la plateforme de signalisation du point de contrôle du fuseau (SAC) portée par KNL1 avec les modules d’attachement microtubulaire, assurant une coordination étroite entre mécanique du kinétochore et signalisation.
Multiples interactions de MIS12C avec KNL1
(Figure : vetopsy.fr d'après Yatskevich et coll)
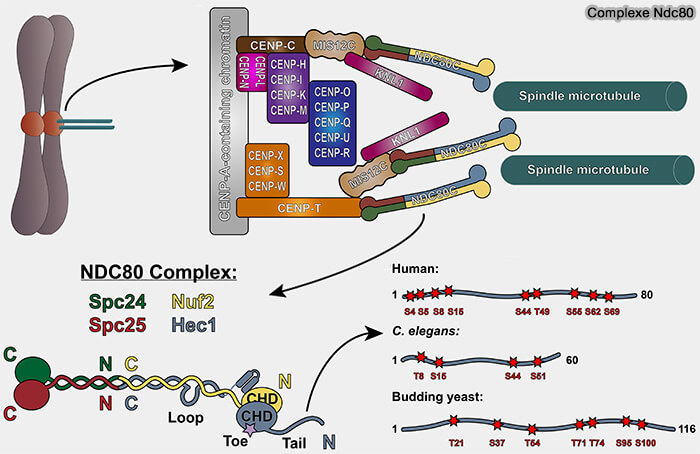
Le complexe NDC80 constitue la principale structure d’attachement des microtubules au kinétochore.
1. Ce complexe est formé par :
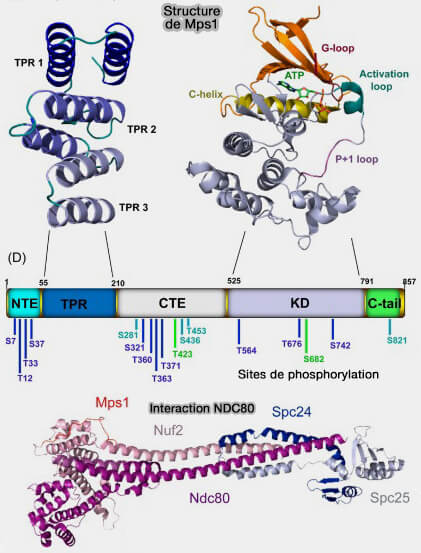
MPS1 et interaction avec NDC80
(Figure : vetopsy.fr d'après Bolanos-Garcia) NDC80 (Hec1) et NUF2 qui s’associent pour former une longue tige en coiled-coil terminée par un domaine CH capable de reconnaître et de se lier à l'αβ-tubuline des microtubules.
SPC24 et SPC25 qui forment le domaine basal du complexe s’ancrant sur le complexe Mis12 et assurant la connexion avec la couche interne.
2. Au-delà de son rôle mécanique, le complexe NDC80 joue un rôle clé dans la signalisation en couplant l’état d’attachement microtubulaire à l’activation ou l’inhibition du point de contrôle du fuseau (SAC).
Lorsqu’un microtubule s’attache correctement, ce domaine est occupé par la tubuline, empêchant le recrutement de MPS1, i.e. cette exclusion constitue un signal local de levée du SAC sur les kinétochores attachés.
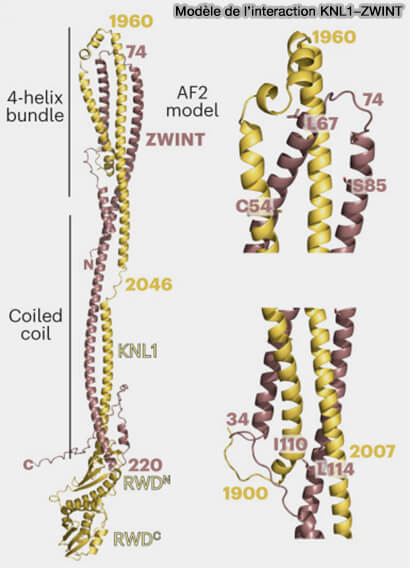
KNL1
KNL1 (Kinetochore Null 1), également appelée CASC5 chez les mammifères, est une grande protéine d’échafaudage localisée à la base du réseau KMN.
Structure de KNL1
1. KNL1 est majoritairement intrinsèquement désordonnée, caractéristique des protéines d’échafaudage du kinétochore externe.
Interaction KNL1-ZWINT
(Figure : vetopsy.fr d'après Polley et coll)
Elle s’insère dans le réseau KMN par une interaction stable avec le complexe Mis12, qui assure son positionnement précis à la base de la couche externe du kinétochore.
Cette organisation confère à KNL1 une géométrie flexible, lui permettant d’exposer sa région N-terminale vers l’espace cytoplasmique tout en restant ancrée à la plateforme centromérique.
Par cette disposition, KNL1 occupe une position stratégique adjacente au complexe Ndc80, sans interagir directement avec lui, et se trouve idéalement placée pour coordonner les événements mécaniques et signalétiques (SAC) au niveau du kinétochore.
2. Le domaine C-terminal de KNL1 contient deux domaines RWD qui établissent des interfaces distinctes avec le complexe Mis12C.
Le domaine RWD-C reconnaît une surface formée par Dsn1 et Pmf1 dans Mis12C,
Le domaine RWD-N se lie à la queue C-terminale de NSL1, consolidant ainsi l’intégration de KNL1 dans le réseau KMN.
En l’absence d’attachement microtubulaire, la kinase MPS1 accède à ces motifs et les phosphoryle, déclenchant le SAC.
Lorsque l’attachement aux microtubules est établi et que la tension est générée, la déphosphorylation des motifs MELT par les phosphatases PP1 et PP2A-B56 entraîne la dissociation progressive des composants du SAC, conduisant à sa levée.
Remarque : certains facteurs non constitutifs, impliqués dans le trafic membranaire comme SNAP29, peuvent perturber indirectement l’assemblage fonctionnel du kinétochore, notamment le recrutement de KNL1.





 différences structurales avec H3).
différences structurales avec H3).