La terminaison de la réplication met fin à l’activité du réplisome tandis que la correction d’épreuve des polymérases réplicatives élimine immédiatement les nucléotides mal incorporés et assure la fidélité de la synthèse de l’ADN.
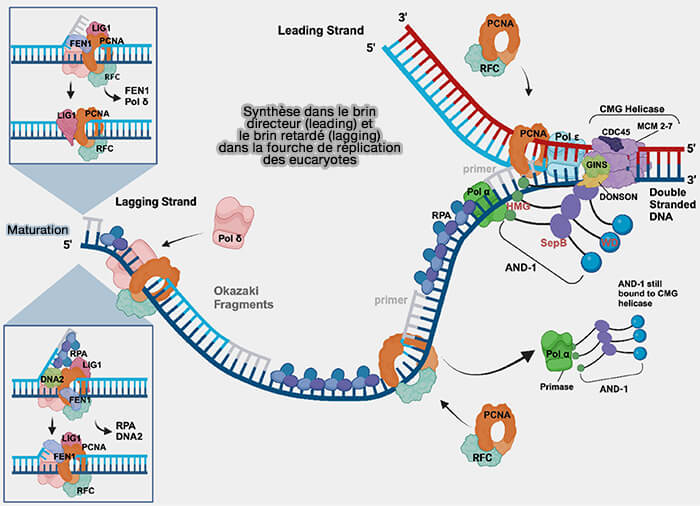
Synthèse dans le brin directeur et retardé de la fourche de réplication eucaryote
(Figure : vetopsy.fr d'après Nasheuer et coll)
Terminaison de la réplication de l’ADN
Mécanisme de terminaison des fourches de réplication
1. À mesure que la réplication progresse, les fourches issues d’origines voisines avancent l’une vers l’autre et la région d’ADN parental située entre elles devient progressivement de plus en plus courte.
Les deux polymérases terminent la copie des dernières bases du brin matrice et synthétisent les derniers fragments d’Okazaki, les brins néosynthétisés arrivent bord à bord.
La maturation des fragments d’Okazaki assure la jonction progressive de chaque fragment avec le fragment adjacent de sorte que les segments d’ADN synthétisés par les deux fourches deviennent continus et se rejoignent sans qu’il subsiste d’ADN parental non copié.
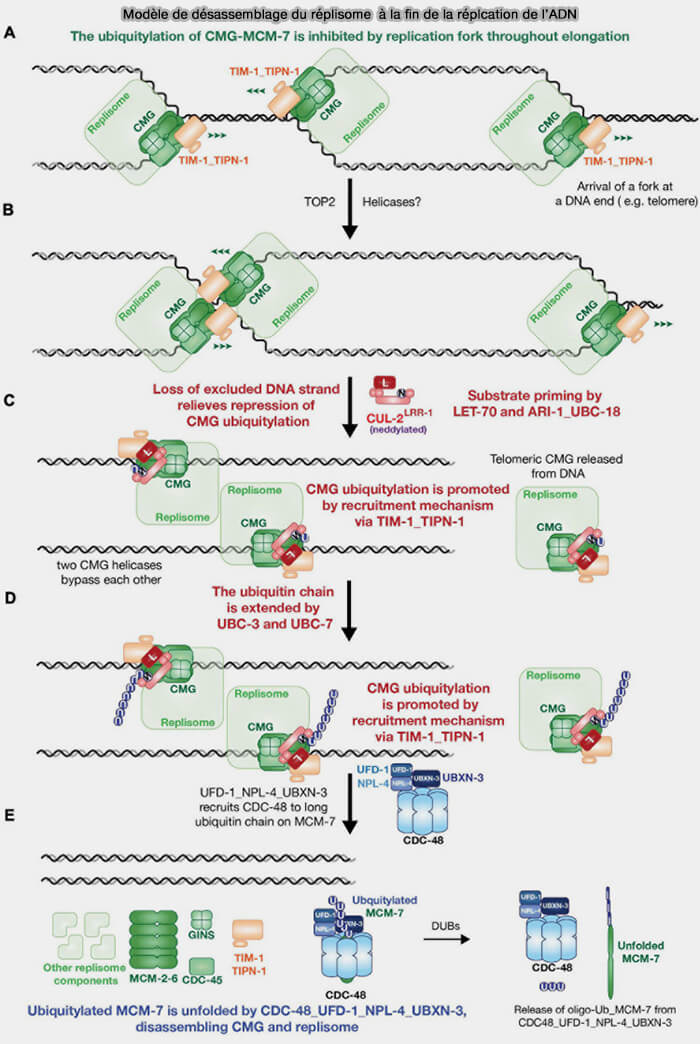
Désassemblage du réplisome
(Figure : vetopsy.fr d'après Xia)
3. Quand les deux fourches convergentes ont achevé la synthèse et que l’ADN situé entre elles a été entièrement dupliqué, l’hélicase CMG (Cdc45-MCM2-7-GINS), qui progressait jusque-là sur l’ADN simple brin du brin directeur matrice, se retrouve désormais encerclant de l’ADN double brin nouvellement formé, substrat sur lequel elle ne peut plus poursuivre sa translocation.
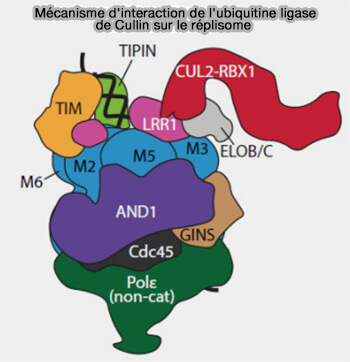
Interaction ubiquitine ligase avec le réplisome
(Figure : vetopsy.fr d'après Jones et coll)
Chez les métazoaires, la première étape de ce processus correspond à l’ubiquitination spécifique de la sous-unité MCM7 du complexe CMG par la cullin-RING ligase CRL2Lrr1, modification qui marque l’hélicase pour son déchargement de la chromatine.
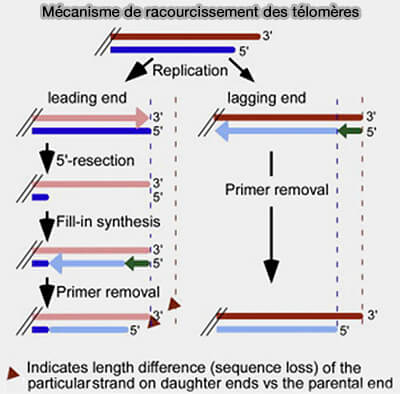
Mécanisme de raccourcissement des télomères
(Figure : vetopsy.fr d'après Wellinger)
a. Lorsque l’amorce d’ARN terminale est éliminée, il n’existe plus de fragment d’Okazaki en aval pour fournir une extrémité 3'-OH permettant l’extension par une ADN polymérase.
La polymérase ne peut donc pas combler cette région.
Il reste alors une extrémité simple brin du côté du brin matrice, appelée overhang télomérique.
b. Il en résulte une perte progressive de séquence à l’extrémité des chromosomes à chaque cycle de réplication, phénomène connu sous le nom de problème de l’extrémité réplicative.
2. Sur le brin directeur, la réplication peut théoriquement se poursuivre jusqu’à l’extrémité du chromosome et produire initialement une extrémité double brin franche (blunt).
Une extrémité blunt correspond à une extrémité où les deux brins d’ADN se terminent au même nucléotide,
a. Toutefois, les télomères fonctionnels possèdent normalement un surplomb simple brin 3' riche en G.
Après la réplication, l’extrémité du brin directeur est donc traitée enzymatiquement.
Le brin complémentaire riche en C est résecté en 5' par des nucléases, notamment Apollo (recrutée par TRF2) puis EXO1, ce qui génère progressivement le surplomb télomérique 3' caractéristique.
b. Ce traitement post-réplicatif convertit l’extrémité blunt produite par la réplication en la structure télomérique fonctionnelle présente aux extrémités des chromosomes.
3. Les télomères (séquences répétées TTAGGG chez l’humain) constituent une solution évolutive qui sert de zones tampons non codantes, leur raccourcissement n’affecte pas immédiatement les gènes.
Les histones H3 et H4 parentales peuvent être recyclées et redistribuées sur les deux molécules d’ADN filles, tandis que de nouvelles histones sont incorporées pour compléter les nucléosomes.
b. Ce transfert est assuré par des chaperonnes d’histones associées à la fourche de réplication.
La sous-unité MCM2 du complexe hélicase MCM2-7 possède un domaine de liaison aux histones capable de capturer les histones parentales H3-H4 au moment du démantèlement du nucléosome et de favoriser leur redistribution sur l’ADN fils.
La chaperonne ASF1 (Anti-silencing function 1) interagit avec les dimères H3-H4 et participe à leur transfert vers les complexes d’assemblage de la chromatine.
c. Ce recyclage contribue à la transmission des marques épigénétiques présentes sur les histones parentales.
Désassemblage et réassemblage des nucléosomes couplés à la réplication
(Figure : vetopsy.fr d'après Rowlands et coll)
3. Les nucléosomes ne peuvent cependant pas être reconstitués uniquement avec les histones recyclées et des histones nouvellement synthétisées doivent être également incorporées (dimérisation et assemblage des histones).
a. Le complexe CAF-1 (Chromatin Assembly Factor 1) joue un rôle central dans ce processus.
Cette chaperonne d’histones interagit directement avec le clamp réplicatif PCNA associé à l’ADN polymérase, ce qui la positionne immédiatement derrière la polymérase et permet de coupler étroitement l’assemblage des nucléosomes à la progression de la réplication.
CAF-1 dépose des dimères d’histones H3-H4 néosynthétisées sur l’ADN nouvellement répliqué, permettant la formation du tétramère (H3-H4)2 qui constitue le cœur du nucléosome.
Une fois ce tétramère formé, NAP1 et des chaperonnes apparentées ajoutent les deux dimères H2A-H2B, complétant ainsi l’octamère histonique du nucléosome.
b. D’autres chaperonnes, notamment FACT (Facilitates Chromatin Transcription), participent également à la dynamique des nucléosomes au niveau de la fourche.
FACT déstabilise transitoirement les nucléosomes en retirant ou en relocalisant un dimère H2A-H2B, ce qui permet aux complexes enzymatiques volumineux (réplisome ou ARN polymérase) de traverser la chromatine, tout en limitant la perte d’histones.
Après le passage de la fourche, FACT facilite la réassociation des dimères H2A-H2B avec le tétramère H3-H4 restant.
FACT agit donc principalement comme chaperonne de dynamique et de stabilisation des nucléosomes, plutôt que comme chaperonne principale de dépôt.
Remarque : FACT intervient également lors de l’élongation transcriptionnelle, où ce complexe facilite le passage de l’ARN polymérase II à travers les nucléosomes en déstabilisant transitoirement certaines interactions histone-ADN puis en favorisant le réassemblage local de la chromatine derrière l’enzyme.
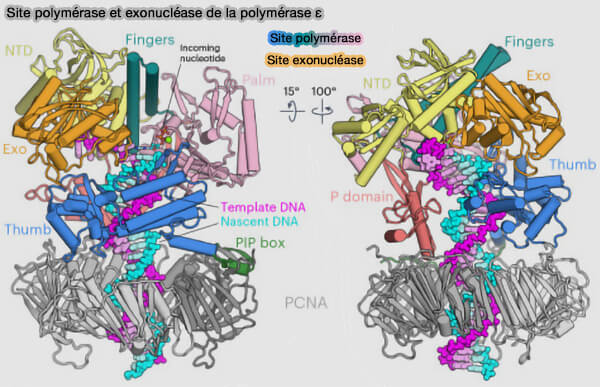
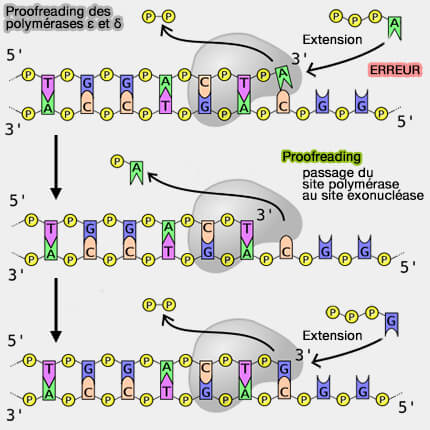
Correction d’épreuve (proofreading) lors de la réplication
1. La fidélité de la réplication de l'ADN résulte ainsi de plusieurs niveaux de contrôle :
Les polymérases réplicatives ε et δ disposent d'un site polymérase (synthèse 5' → 3'), mais aussi d’un site exonucléase 3' → 5', séparé d’environ 30-40 Å, qui permet d’éliminer les nucléotides incorrectement incorporés au cours de l’élongation.
L'efficacité du proofreading est remarquable car le taux d’erreur brut de la polymérase ≈ 10-5, après proofreading ≈ 10⁻-7, après réparation des mésappariements ≈ 10-9-10-10.
Sites polymérase et exonucléase de la polymérase ε
(Figure : vetopsy.fr d'après Roske et Yeeles)
2. La correction d’épreuve agit immédiatement après l’incorporation d’un nucléotide incorrect et se déroule donc pendant l’élongation du brin d’ADN, bien que ce mécanisme soit présenté à la fin du chapitre consacré à la réplication pour expliquer la fidélité globale du processus.
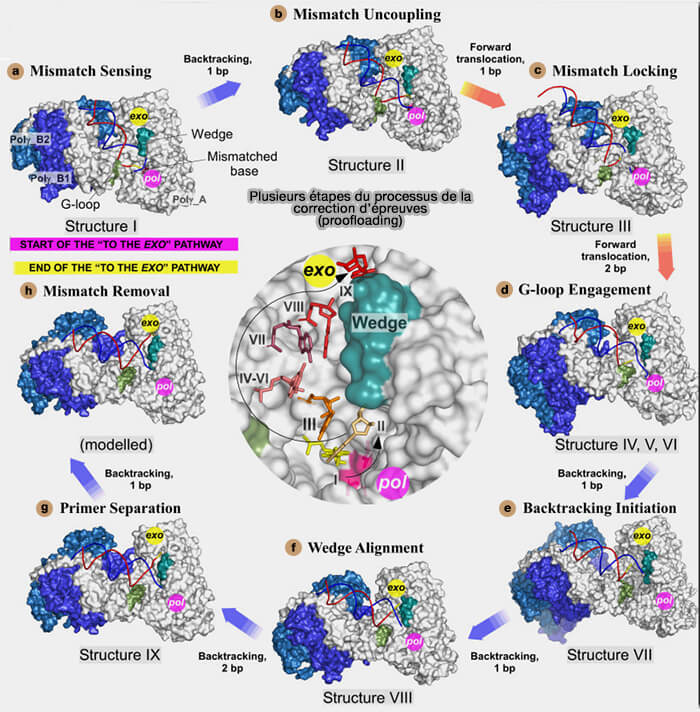
3. Le mécanisme de réparation du mauvais appariement de bases (mismatch), expliqué par la figure ci-dessous, est le suivant (Structural basis for DNA proofreading 2023).
a. Le brin bleu (matrice) est correctement positionné dans la gorge de la polymérase, le brin rouge forme un ADN double brin (dsDNA) avec le brin matrice et le nucléotide jaune est à l’extrémité 3' (mismatch sensing).
Cependant ce nucléotide n’est pas correctement apparié avec la base matrice et provoque une distorsion locale du dsDNA.
b. L’extrémité du brin rouge commence à se désapparier et si le nucléotide jaune reste attaché au brin rouge, mais l’appariement avec le brin bleu devient instable (mismatch uncoupling).
c. la polymérase a déjà transloqué vers l’avant, et un nucléotide en position n+1 sert de base de “ verrouillage ” (mismatch locking)
Le mismatch n’est toujours pas excisé, mais la structure est figée dans une configuration qui empêche le retour simple à l’élongation normale et prépare la suite du transfert vers l’exonucléase.
d. La Guide loop ou G-loop entre véritablement en action et participe à la réorientation de l’extrémité du primer (G-loop engagement).
En parallèle, la figure montre une courbure du pouce pendant cette translocation.
Autrement dit, la polymérase n’a pas encore hydrolysé le mauvais nucléotide, mais elle réorganise ses éléments structuraux pour engager l’extrémité 3' dans une trajectoire compatible avec le canal exonucléase.
Étapes de la correction d'épreuves (proofloading)
(Figure : vetopsy.fr d'après Buchel et coll)
e. Après cette translocation avant, l’enzyme commence une translocation arrière (backtracking initiation).
La polymérase commence à “ reculer ” par rapport à l’ADN, ce qui aide à repositionner le primer et à rapprocher le mismatch de l’entrée du canal exonucléase.
Ce n’est donc pas un simple glissement passif du brin, mais une réorganisation mécanique coordonnée enzyme-ADN.
f. Le mismatch arrive à l’entrée du canal exonucléase, et la wedge est correctement alignée pour favoriser la séparation terminale du primer (wedge alignment).
La paire mal appariée est visible à l’entrée du canal exo.
Le mauvais nucléotide n’est pas encore totalement dans le site exonucléase, mais il est maintenant orienté vers lui.
Remarque : dans ce contexte, wedge signifie littéralement “ coin structural ”, élément protéique de la polymérase qui aide à écarter localement l’extrémité de l'amorce de l'ADN double brin (dsDNA).
Son rôle n’est pas de couper l’ADN, mais de favoriser la séparation terminale nécessaire pour que l’extrémité 3' quitte la trajectoire polymérase et soit orientée vers le site exonucléase.
g. L’extrémité 3' de l'amorce est finalement positionnée dans le site exonucléase (primer separation).
Autrement dit, le brin rouge est maintenant suffisamment séparé du brin bleu pour que l’extrémité terminale portant le mauvais nucléotide jaune puisse être prise en charge par le centre exonucléolytique.
C’est l’aboutissement du transfert du site polymérase vers le site exonucléase.
Remarque : le fraying désigne le désappariement progressif de l’extrémité de l'ADN double brin (dsDNA) et correspond à une phase de transition qui commence après la détection du mismatch et se poursuit jusqu’à la séparation suffisante du primer.
h. Enfin, le nucléotide incorrect jaune est excisé par hydrolyse dans le site exonucléase (mismatch removal).
Une fois le nucléotide fautif éliminé, l’extrémité 3'-OH correcte est ensuite repositionnée dans le site polymérase, ce qui permet la reprise immédiate de la synthèse avec l’incorporation du nucléotide approprié.
Les erreurs de réplication qui échappent à la correction d’épreuve sont éliminées par les systèmes de réparation de l’ADN, notamment la réparation des mésappariements (MMR), qui sont étudiés dans le chapitre consacré à la réparation de l’ADN.





 maintien des télomères par la télomérase).
maintien des télomères par la télomérase). 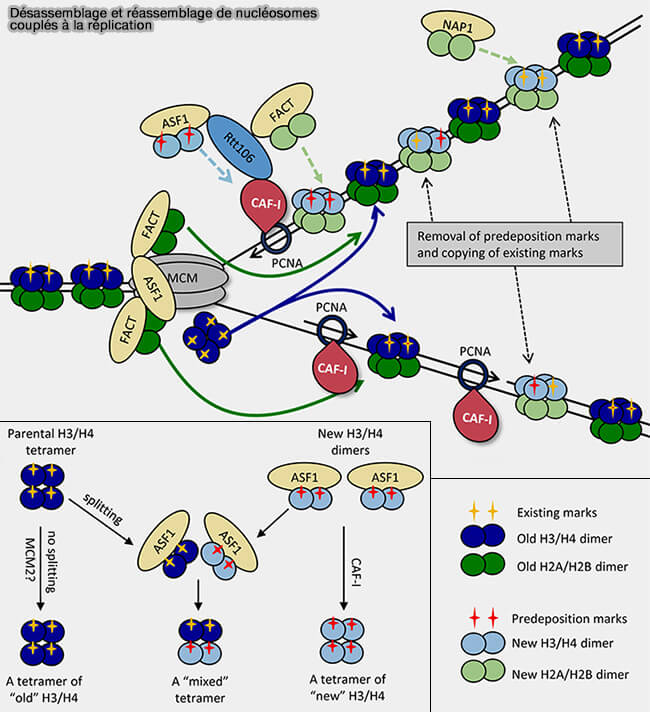
){kind=link}