


Acides nucléiques
Chromatine
Chromatosome et fibre de 30 nm
- Biochimie
- Chimie organique
- Bioénergétique
- Composition de la matière vivante
- Glucides
- Lipides
- Coenzymes
- Hormones
- Composés inorganiques

Le chromatosome, formé du nucléosome associé à l’ADN de liaison et à l’histone H1, constitue une étape de compaction intermédiaire dont l’organisation a été décrite par le modèle historique de la fibre de 30 nm.
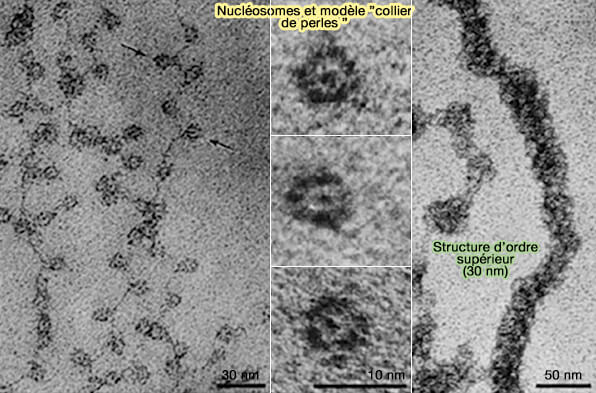
(Figure : vetopsy.fr d'après Mattick et Amaral)
Chromatosome
Le nucléosome, l'ADN de liaison et l'histone H1 constituent le chromatosome.
ADN de liaison
L'ADN de liaison (linker DNA) est le double brin d'ADN qui sépare deux nucléosomes adjacents, correspondant aux segments d’ADN entrant et sortant du nucléosome, dont la longueur varie selon les espèces et le type de cellule.
Chez les mammifères, il contient généralement entre 20 et 80 paires de bases (pb), i.e. 10-50 chez la levure.
1. Cet ADN est connecté à :
- à l'histone H1 qui lui-même est attaché au nucléosome,
- à H2A dont la queue C-terminale peut interagir avec l’ADN de liaison, contribuant à la stabilisation locale et pouvant introduire une asymétrie structurale du chromatosome (Distinct Structures and Dynamics of Chromatosomes with Different Human Linker Histone Isoforms 2021).
2. L’ADN de liaison constitue la région la plus accessible de la fibre de chromatine et est particulièrement sensible à l’action de nucléases, qui produisent d’abord des unités chromatosomiques isolées, puis, si la digestion se poursuit, des nucléosomes dépourvus de l’ADN d’entrée/sortie stabilisé par H1.
Cette propriété a été mise en évidence expérimentalement par digestion de la chromatine avec des enzymes comme la micrococcal nuclease (MNase) ou la DNase I, qui clivent préférentiellement l’ADN linker entre les nucléosomes.
Remarque : les cellules possèdent plusieurs désoxyribonucléases endogènes impliquées dans la dégradation de l’ADN au cours de processus physiologiques tels que :
- DNase I dans l’apoptose et dans certains mécanismes de remodelage ou de dégradation extracellulaire de l’ADN,
- DNase II dans la digestion lysosomale de matériel nucléaire.
Ces enzymes peuvent cliver préférentiellement les régions d’ADN les plus accessibles, comme l’ADN de liaison entre les nucléosomes.
3. La longueur et la conformation de l’ADN de liaison déterminent :
- la distance inter-nucléosomique,
- l’angle entre nucléosomes adjacents,
- la propension à former des fibres de chromatine plus ou moins compactes, plutôt qu’une fibre régulière unique.
Histone H1
L’histone H1 n’est pas une composante du cœur du nucléosome et est présente en une seule copie par nucléosome, contrairement aux autres histones.
Moins conservée que les histones du cœur du nucléosome (H2A, H2B, H3 et H4), l’histone H1 présente une plus grande variabilité de séquence et plusieurs variants selon les tissus et les stades du développement.
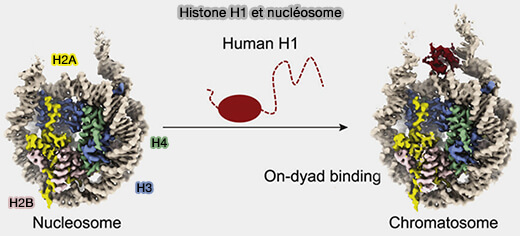
(Figure : vetopsy.fr d'après Zhou et coll)
Une grande partie des queues N- et C-terminales sont intrinsèquement désordonnées, ce qui rend difficile la résolution de la structure complète de l’histone H1 associée au nucléosome, et, de plus, il existe différents variants ( histone H1).
histone H1).
1. Toutefois, de manière générale, H1 possède un domaine globulaire central qui se fixe au niveau de l’ADN de liaison en deux endroits, i.e. à l’entrée/sortie de l’ADN du nucléosome.
- Le domaine globulaire de H1 reconnaît le squelette phosphate et le sillon mineur, stabilisant l’ADN sur l’octamère.
- Ses extrémités N-terminale et C-terminale, riches en lysine, interagissent de façon électrostatique avec l’ADN de liaison, mais aussi avec les nucléosomes adjacents, notamment via des contacts avec l’ADN et les surfaces histoniques exposées, favorisant la compaction en fibre chromatinienne.
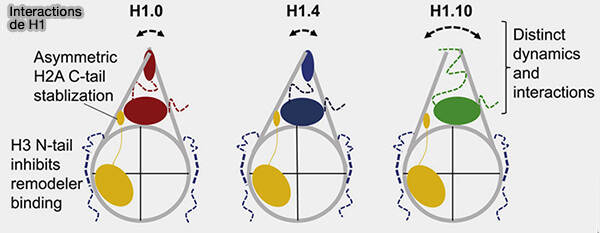
(Figure : vetopsy.fr d'après Zhou et coll)
2. Son rôle est de :
- stabiliser la particule nucléosomique en stabilisant l’angle entre les brins d’ADN entrant et sortant du nucléosome,
- favoriser des états de compaction plus élevés de la chromatine, historiquement associés à la fibre de 30 nm, mais correspondant en réalité à des organisations dynamiques et hétérogènes,
- limiter l’accessibilité de l’ADN aux facteurs de transcription.
Fibres de 30 nm
Structure
La fibre de 30 nm de diamètre est une structure filamenteuse, constituée d’un enchaînement de nucléosomes organisés de manière régulière, historiquement décrite comme un niveau supérieur de compaction de la chromatine.
Cette fibre est classiquement associée à la présence de l’histone H1, qui se fixe sur l’ADN de liaison et peut favoriser des compactions de la chromatine plus élevées.
1. Cette compaction intermédiaire entre nucléosome et chromosome a été proposée comme modèle d’organisation permettant de réduire la longueur de l’ADN tout en conservant une accessibilité modulable pour les processus nucléaires, i.e. transcription, réplication, réparation.
2. En fait, in vivo, la chromatine apparaît plutôt sous forme de polymère chromatinien désordonné et dynamique ou de domaines plus hétérogènes.

Dans les cellules vivantes, une fibre de 30 nm parfaitement organisée est rarement observée, voire absente selon de nombreuses études, et reste essentiellement un modèle conceptuel basé sur des expériences réalisées in vitro.
Modèles géométriques
Bien que la fibre de 30 nm ait été proposée comme un niveau intermédiaire de compaction de la chromatine, les modèles géométriques associés (solénoïde et zig-zag), issus principalement d’études in vitro, ne reflètent pas nécessairement l’organisation réelle in vivo.
Ils correspondent plutôt à des états locaux ou transitoires de compaction, dérivés des interactions entre nucléosomes, pouvant s’observer dans certaines conditions expérimentales et s’inscrivant dans un continuum d’organisations intermédiaires de la chromatine (New insights into nucleosome and chromatin structure: an ordered state or a disordered affair? 2013).
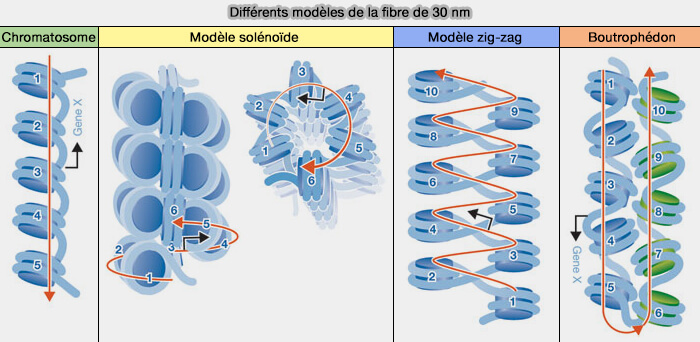
(Figure : vetopsy.fr d'après Quénet et coll)
1. Le modèle en solénoïde, dit à une entrée, est un modèle dans lequel les nucléosomes s’enroulent en hélice, avec environ 6 nucléosomes par tour avec un pas de 11 nm (Single-molecule force spectroscopy reveals a highly compliant helical folding for the 30-nm chromatin fiber 2007).
Le modèle a été conçu par l'observation en microscopie électronique sur des nucléofilaments de longue taille et contenant l'histone H1.
a. Dans ce modèle, l’ADN de liaison est plié de telle sorte qu’il ramène le fil nucléosomique vers le nucléosome suivant le plus proche, situé juste à côté dans l’hélice.
- Géométriquement, chaque nucléosome est donc connecté à son voisin immédiat, avec une seule trajectoire d’entrée-sortie de l’ADN.
- Le cheminement est continu et circulaire, comme un ressort enroulé, i.e. on parle alors de fibre à une entrée.
(Figure : vetopsy.fr modifiée d'après MacGinty et Tan et Kalashnikova et coll)
b. Dans le modèle solénoïde, la compaction de la fibre de 30 nm nécessite non seulement l’empilement des nucléosomes, mais aussi des interactions spécifiques entre histones.
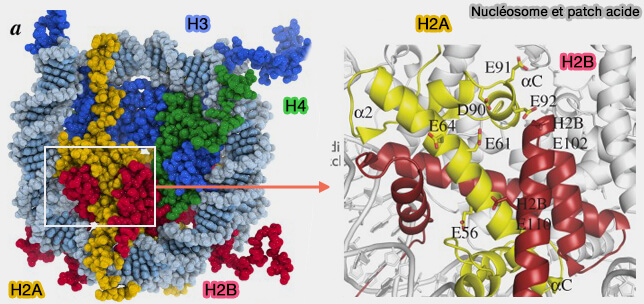
- Les histones H2A et H2B possèdent à leur surface des zones chargées négativement, appelées patch acide (The role of the nucleosome acidic patch in modulating higher order chromatin structure 2013).
- Ces patchs peuvent former des contacts électrostatiques avec les queues N-terminales des histones H4 des nucléosomes voisins.
- Ces interactions stabilisent l’organisation en hélice du solénoïde, en alignant les nucléosomes autour d’un axe central.

Les queues N-terminales d’H4 jouent un rôle de pont, reliant les patchs acides et assurant la cohésion de la fibre.
c. Les modifications sur la queue d'H4 peuvent déterminer le compactage et donc, fournir un niveau de régulation.
Par exemple, l’acétylation sur la lysine16 (H4K16ac) peut arrêter la formation du solénoïde.
2. Le modèle en zig-zag, dit à deux entrées, suppose que les nucléosomes alternent en deux rangées, reliés par l’ADN de liaison tendu, les nucléosomes étant les uns au-dessus des autres.
Le modèle a été conçu par structure cristallographique de 4 et 12 nucléosomes sans H1.
a. Dans ce modèle, l'ADN est droit, i.e. non plié comme dans le modèle précédent.
- Quand l’ADN sort du nucléosome, son voisin le plus proche géométriquement est placé sur la même rangée, et l’ADN devrait se replier sur lui-même, ce qui est défavorable.
- Au lieu de cela, l’ADN de liaison traverse en ligne droite et rejoint le nucléosome de l'autre rangée.
b Chaque nucléosome est relié à deux voisins situés sur des lignes distinctes, ce qui définit une organisation dite à deux entrées, dans laquelle l'ADN de liaison arrive par deux directions opposées.
3. La différence entre les deux modèles, basés sur des conditions expérimentales différentes, pourrait s’expliquer par la plasticité structurale de la chromatine et la difficulté à distinguer des organisations correspondant à des fibres de 10 nm repliées selon un mode en boustrophédon, i.e. organisation alternée en zig-zag, dans un sens puis dans l’autre (Through thick and thin: the conundrum of chromatin fibre folding in vivo 2012).
Les deux configurations pourraient coexister dans des conditions expérimentales spécifiques plutôt que représenter une organisation universelle in vivo (Evidence for heteromorphic chromatin fibers from analysis of nucleosome interactions 2009).
Compactions supérieures de la chromatine : boucles et domaines
BiochimieChimie organiqueBioénergétiqueAcides nucléiquesNucléotidesADNARNChromatineStructure dynamique de la chromatineNucléosomesHistonesMembres des histonesCode des histonesEuchromatine, hétérochromatine et remodelageChromosomesStructure des chromosomesCentromèreTélomèreProtidesGlucidesLipidesEnzymesCoenzymesVitaminesHormonesComposés inorganiquesTransport membranaireMoteurs moléculairesVoies de signalisation