


Acides nucléiques
ARN messagers (ARNm)
Devenir cytoplasmique
- Biochimie
- Chimie organique
- Bioénergétique
- Composition de la matière vivante
- Transport membranaire
- Moteurs moléculaires
- Voies de signalisation

Le devenir cytoplasmique des ARN messagers résulte d’un équilibre dynamique entre traduction, stockage dans les granules de stress, adressage vers les P-bodies, interférence par l’ARN et dégradation des transcrits.
Principes généraux
Les ARN messagers (ARNm) peuvent être orientés vers différentes voies cytoplasmiques qui déterminent leur devenir fonctionnel, notamment leur traduction, leur stockage transitoire, leur répression traductionnelle ou leur dégradation.
Complexes ribonucléoprotéiques associés aux ARN messagers
Les ARN messagers ne circulent pas librement dans la cellule mais restent associés à de nombreuses protéines formant des complexes ribonucléoprotéiques.
1. Les protéines hnRNP sont associés aux pré-ARNm nucléaires au cours de leur maturation, de leur épissage et de leur préparation à l’export nucléaire.
2. Les particules mRNP (messenger ribonucleoprotein particles) sont des particules ribonucléoprotéiques associant un ARN messager mature à de nombreuses protéines de liaison aux ARN (RBP).
Ces complexes peuvent exister sous différents états fonctionnels au cours de la vie cytoplasmique des ARN messagers :
- les mRNP nouvellement exportées du noyau (
 transport des ARN)
transport des ARN) - les mRNP engagées dans la traduction et associées aux ribosomes,
- les mRNP réprimées dont la traduction est inhibée,
- les mRNP stockées dans les granules de stress,
- les mRNP orientées vers les P-bodies ou vers différentes voies de dégradation cytoplasmique.
Couplage entre traduction et devenir des ARN messagers
Le devenir des ARN messagers cytoplasmiques dépend de nombreux mécanismes capables de modifier la composition des mRNP et d’orienter les transcrits vers la traduction, le stockage, la répression traductionnelle ou la dégradation.
1. L’état de traduction des ARN messagers influence fortement leur stabilité.
- les ARN fortement associés aux ribosomes sont généralement protégés contre certaines voies de dégradation.
- les ARN faiblement traduits ou présentant des anomalies de traduction deviennent plus accessibles aux mécanismes de surveillance des ARN comme la voie NMD ou NGD ( mécanisme de surveillance des ARNm).
2. Les protéines de liaison aux ARN (RBP) peuvent stabiliser ou au contraire déstabiliser les ARN messagers en recrutant différents complexes de dégradation.
Par exemple, les protéines TTP (tristetraprolin) favorisent la dégradation de certains ARNm inflammatoires riches en éléments ARE (AU-rich elements) présents dans leur région 3′UTR (The ARE-binding protein Tristetraprolin (TTP) is a novel target and mediator of calcineurin tumor suppressing function in the skin 2018).
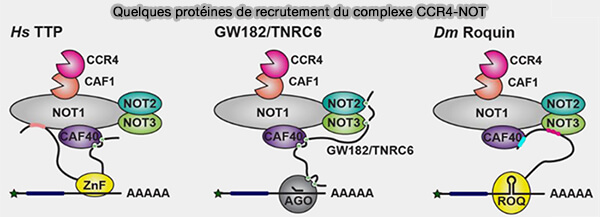
(Figure : vetopsy.fr adaptée d'après Krempl et coll)
3. Certains ARN régulateurs, notamment les miRNA et les siRNA peuvent également orienter les ARN messagers vers des mécanismes de répression traductionnelle et de dégradation cytoplasmique (dégradation dirigée par les ARN régulateurs).
4. Certains signaux cellulaires liés au stress ou à l’état métabolique de la cellule peuvent également modifier le devenir des ARN messagers cytoplasmiques en favorisant leur réorganisation au sein de condensats cytoplasmiques spécialisés associés au stockage, à la répression traductionnelle ou à la dégradation des ARN.

Ces mécanismes permettent de contrôler la durée de vie des ARN messagers et participent ainsi à la régulation de l’expression génique.
Condensats cytoplasmiques associés aux ARNm
1. Les ARN messagers cytoplasmiques ne sont pas uniquement diffusés librement dans le cytosol, mais peuvent également être regroupés au sein de structures ribonucléoprotéiques dynamiques dépourvues de membrane appelées condensats biomoléculaires dont l’assemblage repose notamment sur des interactions multivalentes impliquant des protéines contenant des domaines intrinsèquement désordonnés (IDR, intrinsically disordered regions).
- Les IDR, favorisant la séparation de phase liquide-liquide (LLPS), correspondent à des régions protéiques dépourvues de structure tridimensionnelle stable, capables d’établir de multiples interactions faibles, transitoires et réversibles avec différentes protéines ou ARN.
- Les condensats biomoléculaires résultent notamment du regroupement dynamique de particules mRNP et de nombreuses protéines de liaison aux ARN (RBP), dont la composition varie selon l’état fonctionnel des ARN messagers.
Les condensats moléculaires et la séparation de phase liquide-liquide (LLPS) sont étudiés dans un chapitre spécifique.
2. Parmi ces structures, les corps P (P-bodies) et les granules de stress jouent un rôle majeur dans le stockage, la répression traductionnelle, la surveillance et la dégradation des ARN messagers cytoplasmiques (Mammalian stress granules and P bodies at a glance 2020).
Remarque : d’autres condensats cytoplasmiques associés aux ARN ont également été décrits dans certains contextes cellulaires spécialisés, notamment les P granules/germ granules des cellules germinales, les granules neuronaux impliqués dans le transport local des ARN ou certains condensats associés aux réponses antivirales.

(Figure : vetopsy.fr d'après Sansevrino et coll)
Corps P (P-bodies)
Les corps P (processing bodies ou P-bodies) sont des condensats cytoplasmiques enrichis en facteurs impliqués dans la répression traductionnelle, la surveillance et la dégradation des ARN messagers (ARNm).
Ils constituent des centres dynamiques d’aiguillage des ARN messagers entre traduction, stockage et dégradation.
1. Les corps P contiennent notamment :
- des protéines de décapping comme DCP1, DCP2 et EDC4 impliquées dans l’hydrolyse de la coiffe m7G et l’organisation du complexe de décapping,
- l’exoribonucléase XRN1, assurant la dégradation cytoplasmique des ARN dans le sens 5′ → 3′,
- des protéines associées au complexe CCR4-NOT et au remodelage des complexes ribonucléoprotéiques comme DDX6, PATL1, LSM14 impliquée dans la répression traductionnelle et l’assemblage des P-bodies, ainsi que le complexe LSM1-7,
- des protéines Argonaute et différents composants du complexe RISC impliqués dans les mécanismes d’interférence par l’ARN (RNAi) et la répression traductionnelle médiée par les microARN (miRNA),
- plusieurs facteurs de surveillance des ARN cytoplasmiques, notamment UPF1, SMG5 et SMG7, Pelota et HBS1.
2. Les ARN messagers présents dans les P-bodies sont généralement absents des polysomes (ou polyribosomes) actifs et peuvent :
- être stockés transitoirement,
- retourner vers les polysomes pour reprendre leur traduction,
- être dirigés vers des mécanismes de dégradation cytoplasmique.
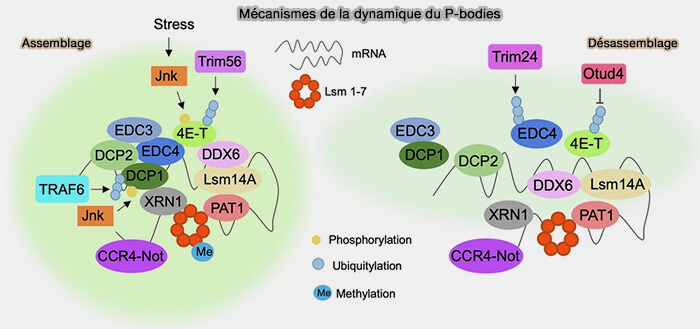
(Figure : vetopsy.fr d'après Senatore et coll)
3. La dynamique d’assemblage et de désassemblage des corps P est également régulée par différentes modifications post-traductionnelles affectant plusieurs composants des condensats ribonucléoprotéiques cytoplasmiques (Reshaping cancer cell plasticity by P-body dynamics and protein translation 2026).
a. L'assemblage est dominé par :
- la kinase JNK (Jun-N-terminal Kinase) qui peut phosphoryler DCP1A et 4E-T lors de différents stress cellulaires, favorisant ainsi l’assemblage des corps P,
- TRIM56, une ubiquitine ligase de type RING, qui module l’ubiquitination de plusieurs composants associés au décapping et aux condensats ribonucléoprotéiques cytoplasmiques,
- la protéine 4E-T (eIF4E-transporter), impliquée dans la répression traductionnelle et le stockage des ARNm, qui participe également à l’organisation des corps P via ses interactions avec DDX6 et EDC4 (A role for the eIF4E-binding protein 4E-T in P-body formation and mRNA decay 2005).
b. À l’inverse, le désassemblage est notamment favorisé par :
- l’ubiquitine ligase TRIM24 qui peut limiter la formation des corps P par ubiquitination d’EDC4, perturbant ainsi l’organisation du complexe de décapping,
- la désubiquitinase OTUD4 qui désubiquitine 4E-T et favorise sa dissociation des corps P, contribuant ainsi à la dynamique de désassemblage de ces condensats ribonucléoprotéiques cytoplasmiques.
Granules de stress et ARN non traduits
Les granules de stress (stress granules) sont des condensats ribonucléoprotéiques cytoplasmiques dynamiques formés en réponse à différents stress cellulaires.

Vous pouvez lire : Stress granules, RNA-binding proteins and polyglutamine diseases: too much aggregation? (2021) qui décrit les mécanismes d’agrégation des protéines de liaison aux ARN, la dynamique des granules de stress et leurs implications dans différentes maladies neurodégénératives.
1. Différents stress cellulaires tels que le stress oxydant, le stress thermique, le stress du réticulum endoplasmique, certaines infections virales ou encore les carences en nutriments activent plusieurs kinases de stress comme HRI, PERK, PKR ou GCN2 capables de phosphoryler la sous-unité eIF2α du facteur d’initiation de la traduction eIF2.
- La phosphorylation de eIF2α inhibe le recyclage du complexe eIF2-GTP-Met-ARNtiMetti, nécessaire à la formation du complexe de pré-initiation traductionnel associé à la sous-unité ribosomique 40S.
- Cette inhibition bloque globalement l’initiation de la traduction, entraîne la dissociation progressive des polysomes (polyribosomes) et favorise l’accumulation d’ARNm non traduits impliqués dans la formation des granules de stress.
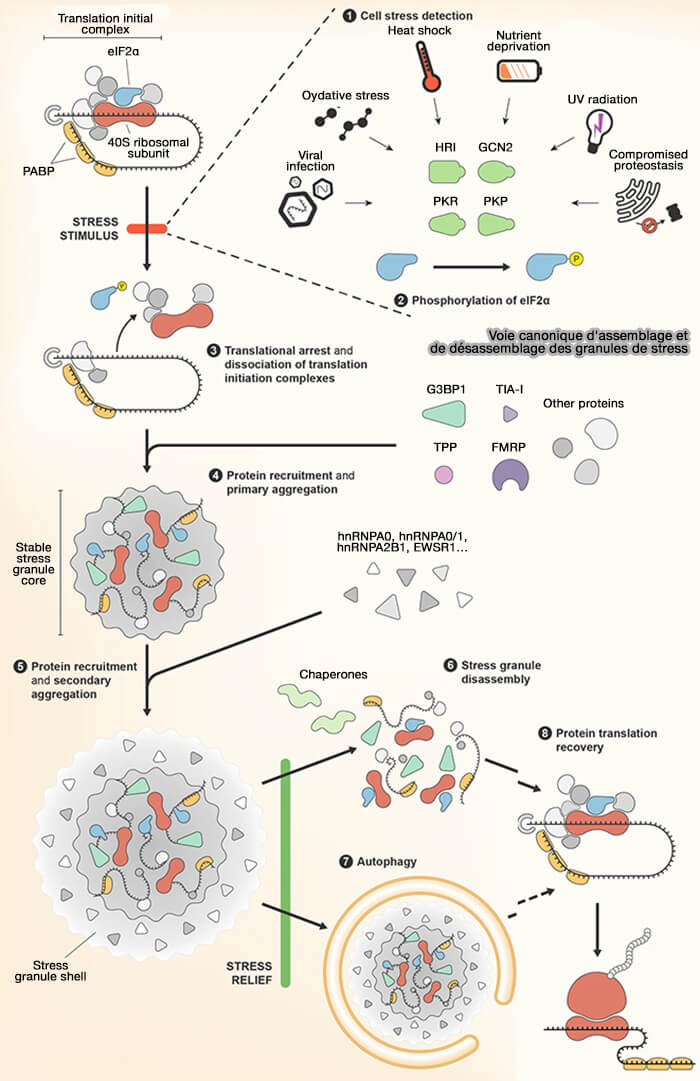
(Figure : vetopsy.fr d'après Marcelo et coll)
2. Le coeur des granules de stress contient principalement (Stress granules, RNA-binding proteins and polyglutamine diseases: too much aggregation? 2021) :
- des ARN messagers (ARNm) dont la traduction a été interrompue,
- des facteurs d’initiation de la traduction,
- des petites sous-unités ribosomiques,
- de nombreuses protéines de liaison aux ARN (RBP).
a. Les principales protéines impliquées dans ces interactions multivalentes comprennent notamment différentes protéines de liaison aux ARN capables de favoriser l’assemblage dynamique de condensats ribonucléoprotéiques cytoplasmiques.
- G3BP1 constitue un facteur central de nucléation des granules de stress en agissant comme plateforme d’assemblage des complexes ribonucléoprotéiques associés aux ARNm non traduits et à différents partenaires protéiques associés aux ARNm réprimés traductionnellement.
- TIA-1 et TIAR, protéines de liaison aux ARN riches en domaines de type prion enrichis en glutamine/asparagine (Q/N-rich domains) favorisent l’agrégation réversible de complexes ribonucléoprotéiques lors de différents stress cellulaires tels que le stress oxydant, le stress thermique ou le stress du réticulum endoplasmique.
- FUS et TDP-43 sont également recrutées dans certains granules de stress et autres condensats ribonucléoprotéiques formés lors de stress cellulaires prolongés ou de perturbations du métabolisme des ARN, notamment dans plusieurs contextes neurodégénératifs.
Leur agrégation pathologique est notamment associée à plusieurs maladies neurodégénératives telles que la sclérose latérale amyotrophique (SLA) et certaines démences frontotemporales.
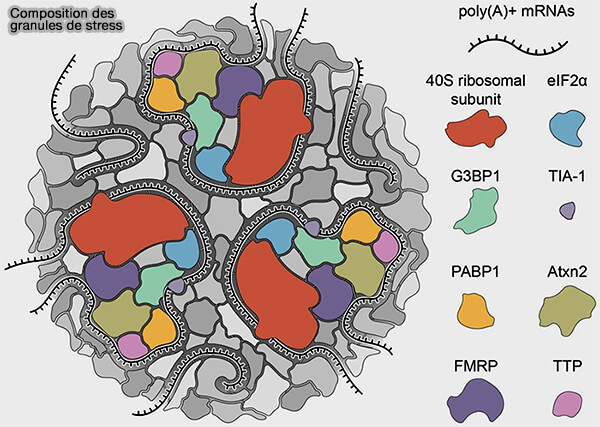
(Figure : vetopsy.fr d'après Marcelo et coll)
b. Autour de ce cœur central s’organise une coque périphérique (shell) plus dynamique contenant différents ARN messagers, protéines de liaison aux ARN (RBP), facteurs de traduction et protéines de signalisation associés de manière plus transitoire aux granules de stress.
4. Les granules de stress constituent des structures dynamiques dont l’évolution dépend de l’intensité et de la durée du stress cellulaire.
- Lors de la résolution du stress, différentes chaperonnes moléculaires et ATPases de remodelage favorisent le désassemblage progressif des granules de stress, permettant la libération des ARN messagers et la reprise de la traduction.
- À l’inverse, lors de stress prolongés ou de défauts de clairance des condensats ribonucléoprotéiques, certains granules de stress peuvent être éliminés par autophagie via des mécanismes parfois désignés sous le terme de granulophagie (The autophagy of stress granules 2023).
5. Ainsi, contrairement aux P-bodies, les granules de stress ne constituent pas principalement des sites de dégradation des ARN mais plutôt des structures transitoires de stockage, de tri et de réorganisation des ARN messagers générant des structures dynamiques capables d’échanger rapidement leurs composants avec le cytoplasme environnant.
- Les granules de stress et les P-bodies interagissent de manière dynamique et peuvent échanger certains ARN messagers ainsi que différentes protéines associées.
- Ces structures participent conjointement au contrôle post-transcriptionnel des ARN cytoplasmiques en fonction de l’état physiologique de la cellule.
Retour aux ARN messagers (ARNm)
BiochimieChimie organiqueBioénergétiqueAcides nucléiquesNucléotidesADNARNStructure de l'ARNARN messager (ARNm) et ARN de transfert (ARNt)ARN ribosomiques (ARNr)ARN régulateurs et fonctionnelsMaturation des ARNÉpissageÉditionÉpitranscriptomeTraductionSurveillance des ARNDégradation des ARNChromatineNucléosomesHistonesChromosomesProtidesGlucidesLipidesEnzymesCoenzymesVitaminesHormonesComposés inorganiquesTransport membranaireMoteurs moléculairesVoies de signalisation