EHD1 lié à l'ATP forme des échafaudages membranaires actifs qui bombent les membranes modèles tubulaires. L'hydrolyse de l'ATP favorise l'auto-assemblage de l'échafaudage, provoquant l'extension du renflement et l'amincissement des régions intermédiaires sur le tubule qui, sur les tubules de moins de 25 nm de rayon, conduit à la scission (ATP-dependent membrane remodeling links EHD1 functions to endocytic recycling 2017).
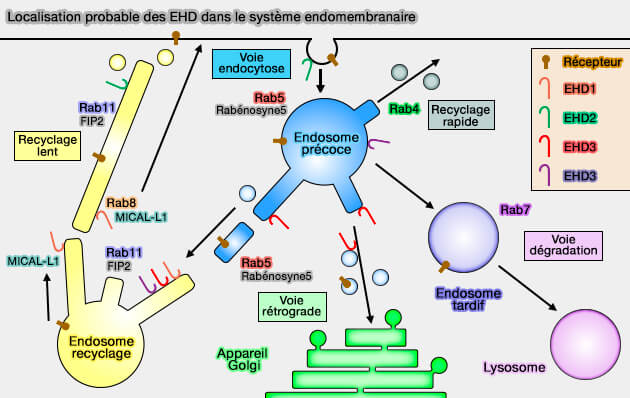
Localisation probable des EHD dans le système endomembranaire
(Figure : vetopsy.fr d'après Naslavsky et coll)
2. EHD3 et EHD4 jouent un rôle dans le transport des endosomes précoces vers les endosomes de recyclage.
EHD1 et EHD3, 86% d'homologie, s'homo- et s'hétérodimérisent et se co-localisent partiellement aux endosomes de recyclage.
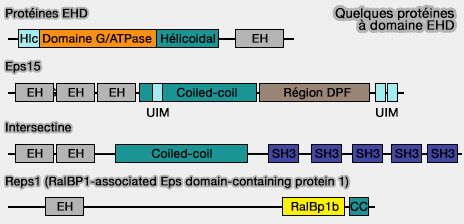
Quelques protéines à domaine EH
(Figure : vetopsy.fr)
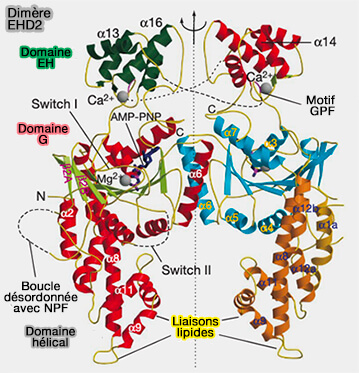
Ce domaine est impliqué dans la dimérisation avec le protomère adjacent et entre dimères ( oligomérisation des EHD).
3. Un domaine charnière est représenté par Pro286 (Pro289 dans EHD4) qui pivote lors de l'activation (Hinge) pour exposer les sites de liaison membranaire, i.e. α9 et α11.
3. Le domaine hélicoïdal, i.e. α8-α12 (287-398), contient les sites de liaison membranaire.
Les dimères EHD2 interagissent avec les membranes via des interactions ioniques médiées par α9-α11 sur une interface incurvée.
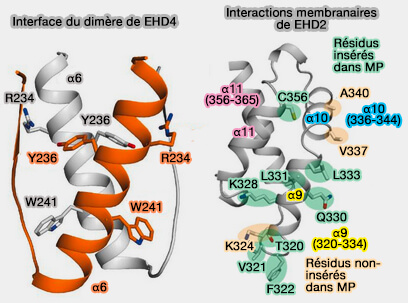
Interactions de EHD4
(Figure : vetopsy.fr d'après Melo et coll)
4. Le lien (400-446) contient le motif GPF qui est impliqué dans l'auto-inhibition du dimère.
5. L'extrémité C-terminale (446-453) est formée par un domaine EH.
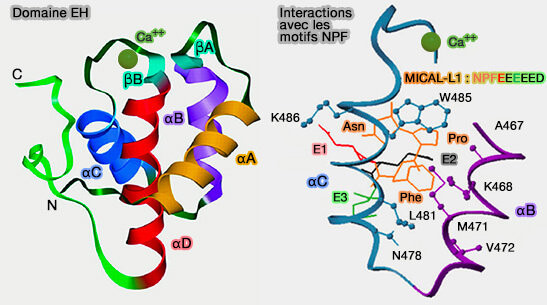
Domaine EH et motifs NPF
1. Le domaine EH est formé par un pli (fold) composé de deux motifs hélice-boucle-hélice étroitement associés, également appelés mains EF (EF-hand), qui peuvent se lier ou non au Ca++ par les hélices αC et αD ( pseudo-mains et mains vestigiales) suivant les protéines EHD, reliées par un court feuillet β antiparallèle.
2. Les protéines contenant des motifs NPF (asparagine-proline-phénylalanine) suivis de résidus acides se lient préférentiellement dans une poche entre les hélices αB et αC du domaine EH, i.e. surface électrostatique chargée positivement ( tableau).
Le résidu +3 par rapport au tryptophane définit la spécificité de la reconnaissance du peptide, i.e. l'alanine ou la sérine ont une préférence pour les NPF, la cystéine ou la valine pour les FW, WW ou SWG.
Pour les motifs NPF, la phénylalanine est le peptide central pour l'ancre hydrophobe, les résidus +1/2/3 par rapport au motif sont essentiels à l'affinité des NPF (par exemple, E comme glutamate pour MICAL-L-1), c'est le cas, en particulier, pour :
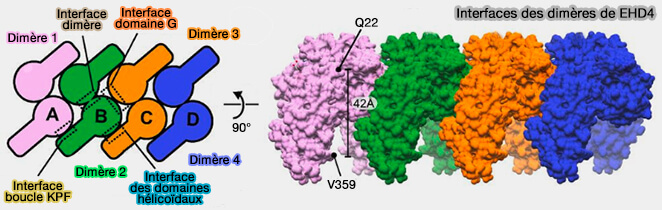
Interfaces des dimères d'EHD4 lors de l'oligomérisation
(Figure : vetopsy.fr d'après Melo et coll)
L'oligomérisation des EHD implique quatre surfaces d'interactions.
1. La première concerne la formation des dimères par l'hélice α6 des deux domaines G (226-245) adjacents.
Les autres sont impliqués dans l'oligomérisation d'ordre supérieur, dimère de dimères.
2. La deuxième implique la boucle KPF (111-134) d'un domaine du dimère 1 avec les hélices α8 et α12 du domaine hélicoïdal du dimère 2.
3. La troisième ferait intervenir les domaines hélicoïdaux comme α8 et α12.
4. La quatrième formerait une interface entre deux domaines G.
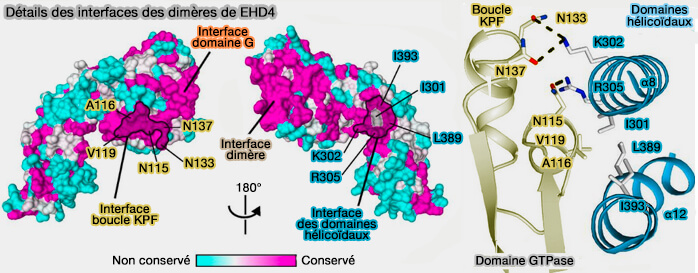
Détails des interfaces des dimères d'EHD4 lors de l'oligomérisation
(Figure : vetopsy.fr d'après Melo et coll)
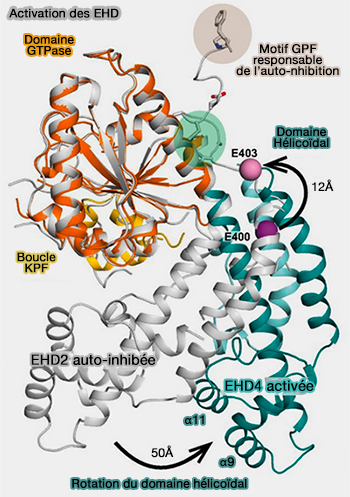
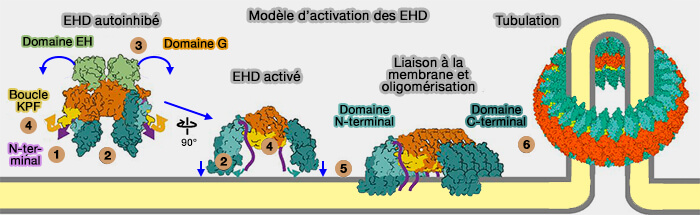
Activation des EHD
Vue d'ensemble
1. En solution, les EHD, comme EHD2, se dimérisent en conformation inhibée.
Les domaines EH se lient à un motif interne Gly-Pro-Phe (GPF) dans le lien entre le domaine hélicoïdal et le domaine EH.
Cette interaction verrouille le domaine EH au domaine GTPase opposé et l'extrémité N-terminale bloque le domaine GTPase, i.e. EHD2 se retrouve en conformation inhibée.
Activation des EHD
(Figure : vetopsy.fr d'après Melo et coll)
2. Lorsque EHD2 est recrutée sur les membranes par sa liaison avec l'ATP, une série de changements conformationnels aligne les sites de liaison des phospholipides avec la membrane et facilite l'oligomérisation de l'EHD2 en structures annulaires (Structural insights into the activation mechanism of dynamin-like EHD ATPases 2017).
EHD2 s'assemble de manière dépendante de l'ATP en oligomères annulaires in vitro.
EHD2 induit la formation de liposomes tubulaires d'un diamètre interne de 20 nm, correspondant au diamètre du col cavéolaire.
Modèle de mécanisme
1. L'activation des EHD est provoquée par la libération de leur extrémité N-terminale du domaine GTPase. L'exemple suivant est proposé pour EHD4.
2. Les domaines hélicoïdaux tournent autour du Pro289 conservé, i.e. un mécanisme identique se retrouve pour la dynamine (power stroke ou torsion du convertisseur).
3. L'extrémité N-terminale des EHD est libérée du domaine GTPase et peut basculer dans la membrane, ajustant ainsi la position des sites de liaison membranaire dans α9 et α11.
Les domaines EH sont déplacés de leur site auto-inhibiteur sur le domaine GTPase.
4. La boucle KPF placée près de l'extrémité N-terminale du domaine GTPase se déplace dans la poche hydrophobe du domaine GTPase.
5. Cette boucle crée une nouvelle interface d'assemblage avec le domaine hélicoïdal du dimère EHD adjacent, stabilisant ainsi la conformation active et favorisant l'oligomérisation des EHD au niveau de la membrane.
La boucle KPF et switch I interagissent de manière dépendante de l'ATP.
Un tel assemblage faciliterait un couplage direct de l'oligomérisation EHD avec la création d'une courbure membranaire.
6. On assiste à la tubulation membranaire.
Modèle d'activation des EHD
(Figure : vetopsy.fr d'après Melo et coll)
L'hydrolyse de l'ATP provoque le désassemblage des oligomères d'EHD qui sont libérés dans le cytosol.
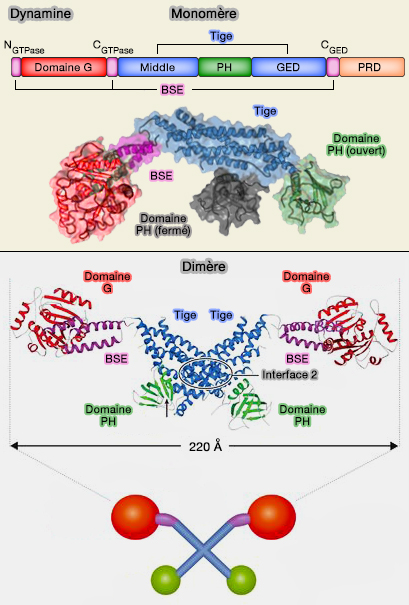
Comparaison avec la dynamine
1. Les modes d'oligomérisation de la dynamine et des EHD diffèrent fondamentalement.
Monomère et dimère de dynamine
(Figure : vetopsy.fr d'après Antonny et coll)
Les domaines GTPase contribuent à l'assemblage en assurant la médiation des contacts dépendants de la GTPase entre les filaments adjacents.
De cette manière, la liaison et l'hydrolyse des nucléotides peuvent induire le réarrangement des filaments adjacents assemblés via la tige, conduisant à une constriction de la membrane sous-jacente.
En revanche, les EHD utilisent une interface unique dans le domaine GTPase pour la dimérisation et utilisent l'interface G dépendante de l'ATP pour une oligomérisation supplémentaire.
Les contacts entre le domaine GTPase et le domaine hélicoïdal du dimère suivant contribuent à la formation d'oligomères.
L'architecture des oligomères EHD exclut un mécanisme de glissement piloté par les nucléotides car les contacts dépendants des nucléotides se forment à l'intérieur et non entre les filaments adjacents.
Cependant, le mode d'assemblage est bien adapté à la stabilisation de la courbure de la membrane par oligomérisation en oligomères tubulaires annulaires.
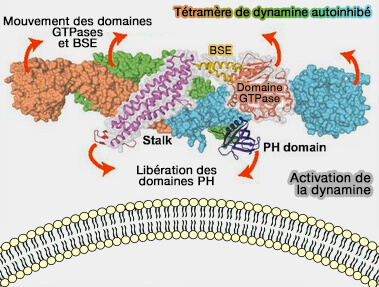
Activation de la dynamine
(Figure : vetopsy.fr d'après Melo et coll)
2. Malgré ces observations, les mécanismes d'activation de la dynamine et des EHD présentent des ressemblances par le levage de l'auto-inhibition.
L'interaction inhibitrice dans la dimérisation des protéines EHD est due au motif Gly-Pro-Phe (GPF) dans le lien entre le domaine hélicoïdal et le domaine EH.
3. De tels contacts inhibiteurs intramoléculaires sont également observés dans d'autres protéines membranaires périphériques ou intégrales.
Les protéines suivantes sont étudiées dans des chapitres spéciaux.
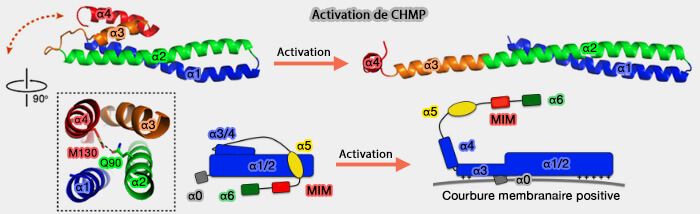
Activation des protéines CHMP
(Figure : vetopsy.fr d'après Tang
et coll)
La formation de nouvelles interactions dans l'oligomère lié à la membrane s'accompagne de la perte d'interactions intramoléculaires dans le dimère auto-inhibé. Un tel mécanisme peut permettre la réversibilité du recrutement membranaire et de l'oligomérisation.




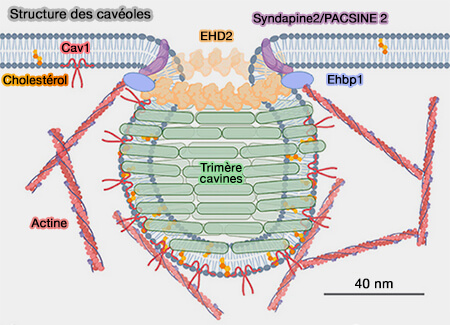
 tableau).
tableau).