L’incorporation de CENP-A correspond au processus d’assemblage de la variante histonique centromérique dans la chromatine, assurant la transmission épigénétique de l’identité du centromère par des mécanismes hautement régulés impliquant HJURP et le complexe Mis18.
Pour préserver l'identité du centromère, les niveaux de CENP-A doivent être maintenus par un mécanisme actif d'incorporation au cours du cycle cellulaire.
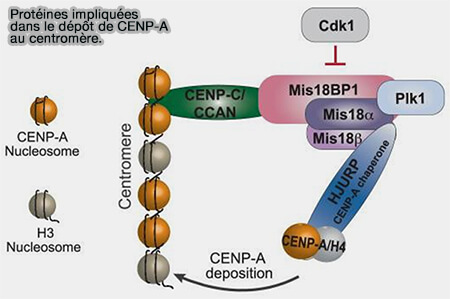
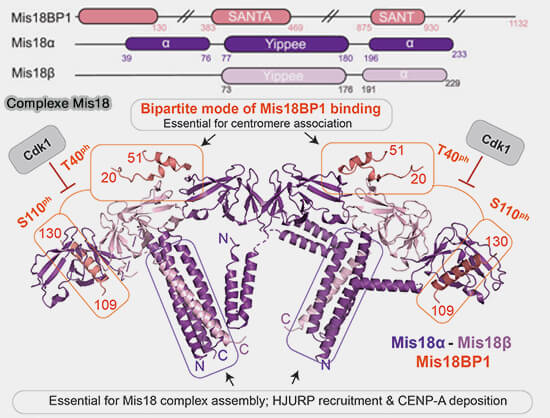
Le complexe Mis18, composé de Mis18α, Mis18β et Mis18BP1 (KNL2), joue un rôle fondamental dans l’assemblage de la chromatine centromérique en amont du dépôt de CENP-A.
Mis18α/β forme un hétéro-hexamère comprenant 4 copies de Mis18α et 2 copies de Mis18β.
Le N-terminal Mis18BP1 interagit avec l’hétéro-hexamère Mis18α/β via l’hétéro-dimère MeDiY pour former un assemblage hétéro-octamérique.
Les sites de phosphorylation consensuels du complexe cycline B/CDK1 (MPF) au sein de Mis18BP1 sont impliqués dans la formation du complexe.
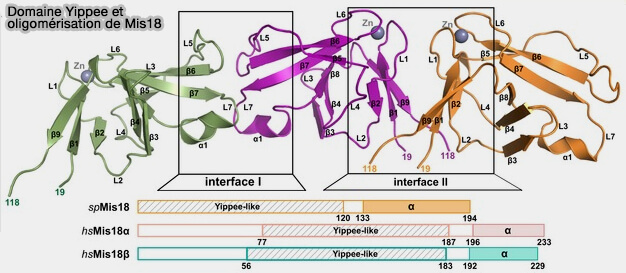
1. Mis18α et Mis18β contiennent tous deux un domaine globulaire appelé domaine Yippee suivi d'une hélice α C-terminale.
Les α-hélices C-terminales de Mis18α et Mis18β forment un triple faisceau hélicoïdal composé de deux Mis18α et un Mis18β. Ensemble, ces régions d'oligomérisation forment un hétéro-hexamère de quatre Mis18α et deux Mis18β.
2. De plus, Mis18α a une région α-hélicoïdale N-terminale supplémentaire qui n'est pas présente dans Mis18β et qui se replie et interagit avec le triple faisceau hélicoïdal formé par les deux α-hélices C-terminales de Mis18α et celle de Mis18β.
3. Mis18BP1, une protéine multi-domaines d'environ 1132 résidus, dont la structure est encore mal connue, comprend :
un domaine N-terminal (0-120) contenant plusieurs sites de phosphorylation, notamment, Thr40 et Ser110 par CDK1 et Thr78 et Ser93 par PLK1 pour réguler ses interactions,
des régions non structurées ou flexibles (linkers), qui permettent des interactions inter-domaines et la flexibilité nécessaire pour s’assembler dans des complexes multi-protéiques.
Interfaces du complexe Mis18
De multiples interfaces inter- et intramoléculaires stabilisent l’architecture tridimensionnelle globale du complexe Mis18 et assurent la localisation du centromère et le fonctionnement de la machinerie de dépôt de CENP-A.
1. L'hétéro-oligomérisation du complexe Mis18α, Mis18β est effectuée via :
a. les modules MeDiY qui comprennent deux copies de Mis18α et une copie de Mis18β.
b. le faisceau hélicoïdal triple C-terminal de de Mis18α/β qui stabilise l’hétéro-oligomérisation en formant le cœur du complexe essentiel pour générer l’assemblage hétéro-octamérique fonctionnel.
Domaines Yippee de Schizosaccharomyces pombe
(Figure : vetopsy.fr d'après Subramanian et coll)
a. Mis18BP1 entre en interaction par deux régions :
un court segment hélicoïdal, résidus 109–130, qui se lie à une poche hydrophobe du domaine Mis18α,
une région, résidus 20-51, contenant deux brins β courts qui interagissent à l'interface Yippee Mis18α/β prolongeant les feuillets à six brins β des domaines Yippee de Mis18α et Mis18β.
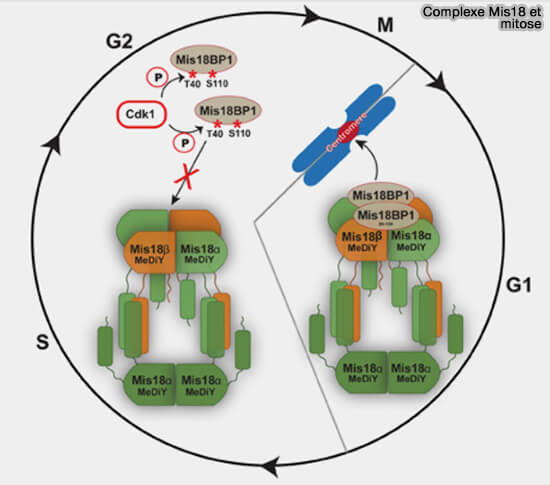
b. Mis18BP1 est phosphorylé sur deux sites de ces régions, Thr40 et Ser110, par le complexe cycline B/CDK1 (MPF), appelé par la suite CDK1.
Remarque : en outre, la phosphorylation de T653 ou T654 de Mis18BP1 par CDK1 est susceptible de perturber l’interaction de Mis18BP1 avec CCAN ( plus loin).
c. L’effet fonctionnel de ces phosphorylations est d’empêcher la formation prématurée du complexe Mis18 pendant la mitose.
La déphosphorylation à la sortie de mitose, lorsque l’activité de CDK1 chute, permet le recrutement de Mis18BP1 sur Mis18α/β, ouvrant la voie au dépôt de CENP-A en début de G1, première phase de l'interphase qui suit la mitose, strictement limité à cette fenêtre temporelle.
Complexe Mis18 et mitose
(Figure : vetopsy.fr d'après Spiller et coll)
Interaction Mis18/centromère
Les mécanismes de recrutement du complexe Mis18 au centromère restent encore débattus.
Ces observations indiquent que Mis18α est recrutée au centromère principalement par l'intermédiaire de Mis18BP1 et que l'intégrité du complexe Mis18 est indispensable à une localisation centromérique efficace.
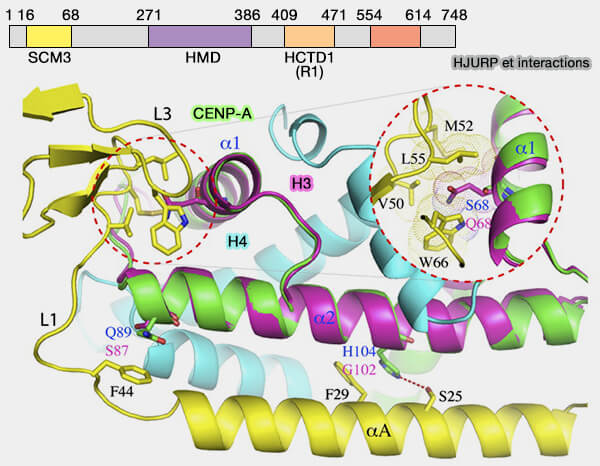
un domaine N-terminal SCM3 (suppressor of chromosome missegregation protein 3), i.e. 16-68, qui se lie à CENP-A,
un domaine intermédiaire central HMD (HJURP middomain), i.e. 271-386 qui se lie directement à l'ADN et participe au chargement de CENP-A aux centromères,
un domaine HCTD1 (HJURP C-terminal domain 1) ou R1, i.e. 409-471, dont la déphosphorylation joue un rôle dans le recrutement de CENP-A,
un domaine HCTD2 ou R2, i.e. 554-614, qui contribue à la dimérisation.
Toutefois, les résidus de CATD enfouis qui génèrent de la rigidité avec H4 sont également nécessaires pour une incorporation efficace dans les centromères.
3. R1 est phosphorylé par CDK1 (Cyclin-dependent kinase 1) sur Ser412, Ser448 et Ser473 pendant la majorité du cycle cellulaire et déphosphorylé à la fin de la mitose.
Elle coopère notamment avec le complexe Mis18, qui prépare la chromatine centromérique à recevoir de nouveaux nucléosomes contenant CENP-A ( interaction HJURP/Mis18).
Interaction HJURP/Mis18
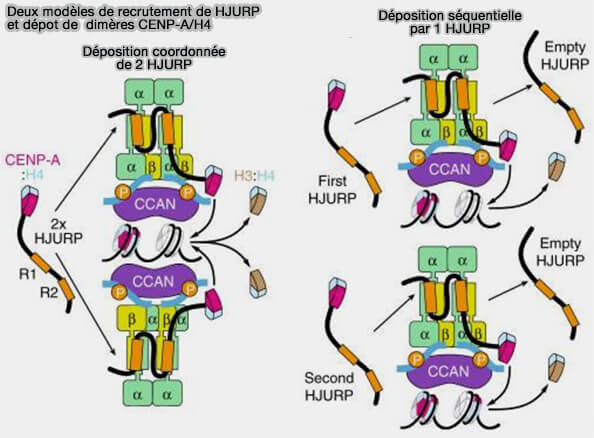
1. Le faisceau hélicoïdal triple C-terminal de Mis18α/β, constitué de deux molécules de Mis18α et d'une molécule de Mis18β, sert de plateforme pour recruter HJURP et les complexes CENP-A/H4.
Mis18α peut se localiser au centromère indépendamment de Mis18β, mais cette dernière est indispensable à un chargement efficace de CENP-A.
Deux modèles de recrutement de HJURP et dépôt de dimères CENP-A/H4
(Figure : vetopsy.fr d'après Pan et coll)
Une fois recrutés au centromère, le complexe Mis18 et HJURP coordonnent l'incorporation de nouveaux nucléosomes contenant CENP-A selon une succession d'événements finement régulés au cours du cycle cellulaire.





 plus loin).
plus loin).