


Acides nucléiques
Chromatine : chromosomes : centromère
CENP-B et liaison spécifique à l’ADN α-satellite
- Biochimie
- Chimie organique
- Bioénergétique
- Composition de la matière vivante
- Glucides
- Lipides
- Coenzymes
- Hormones
- Composés inorganiques

CENP-B est une protéine centromérique qui organise la chromatine des répétitions α-satellites grâce à sa reconnaissance spécifique de la CENP-B box et à ses interactions avec CENP-A et CENP-C, contribuant ainsi à la stabilité du centromère et à l'assemblage du kinétochore.
La reconnaissance spécifique de la CENP-B box par CENP-B la distingue des autres protéines centromériques, qui interagissent principalement avec les nucléosomes CENP-A, ou avec les complexes protéiques du CCAN (Constitutive Centromere-Associated Network),.
- Contrairement à CENP-A, indispensable à l'identité du centromère, CENP-B n'est pas essentielle à la formation d'un centromère fonctionnel (
 néocentromères et chromosome Y).
néocentromères et chromosome Y). - Elle contribue néanmoins au maintien de l'organisation de la chromatine centromérique et au bon fonctionnement du centromère au cours de la division cellulaire.
(Figure : vetopsy.fr)

La CENP-A et le nucléosome CENP-A, spécifiques du centromère, sont étudiés dans un chapitre spécifique.
Structure de CENP-B
CENP-B est une protéine composée de 599 résidus chez l'homme.
1. Elle possède plusieurs domaines fonctionnels distincts.
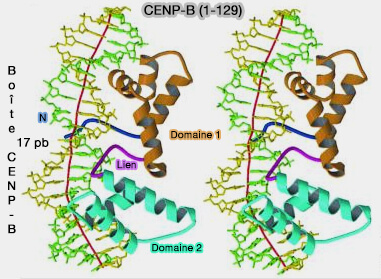
- Le domaine de liaison N-terminal à l’ADN (résidus 1‑129) reconnaît spécifiquement la CENP-B box dans l’ADN α-satellite.
- Le domaine d’oligomérisation C-terminal (résidus 400‑600) permet la formation de dimères, facilitant la structuration de la chromatine centromérique.
- Des régions centrales flexibles connectent les domaines N- et C-terminaux et contribuent à la flexibilité nécessaire pour le pont entre l’ADN et les protéines CENP-A et CENP-C, stabilisant la plateforme protéique du centromère.
2. CENP-B peut former des dimères, ce qui permet de lier simultanément plusieurs boîtes CENP-B et de renforcer la cohésion des unités α-satellites.
Cette capacité d’oligomérisation est cruciale pour l’organisation locale de la chromatine centromérique.
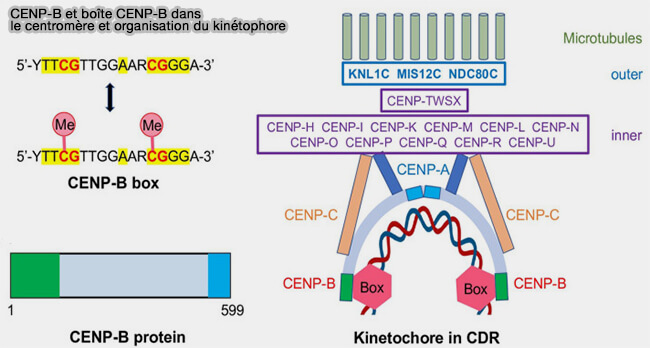
CENP-B et la boîte CENP-B
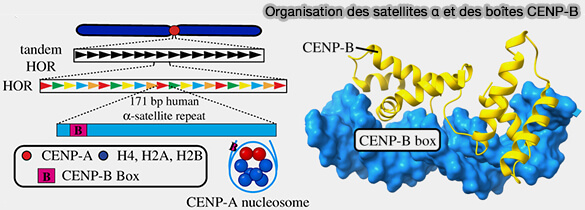
1. Environ la moitié des monomères d'ADN α-satellite contiennent une séquence consensus de 17 paires de bases, appelée boîte CENP-B (CENP-B box), qui constitue le site de liaison spécifique de CENP-B (Preserving centromere identity: right amounts of CENP-A at the right place and time 2025).
(Figure : vetopsy.fr d'après Tanaka et coll)
- La séquence consensus de la boîte CENP-B est 5′-TTCGTTGGAAACGGGA-3′, mais des variations de ce motif sont compatibles avec la liaison de CENP-B.
- Les cytosines présentes dans ce motif peuvent être méthylées, ce qui module la fixation de CENP-B ( méthylation des boîtes).
- L'organisation des boîtes CENP-B au sein des répétitions d'ADN α-satellite présente un motif propre à chaque chromosome, remarquablement conservé entre les haplotypes et les individus malgré la divergence des séquences sous-jacentes (Chromosome-specific centromeric patterns define the centeny map of the human genome 2025).
2. CENP-B se lie aux boîtes CENP-B des répétitions d'ADN α-satellite de la chromatine centromérique grâce à son domaine N-terminal (résidus 1-129), qui comporte deux motifs hélice-tour-hélice (Crystal Structure of the Human Centromere Protein B (CENP-B) Dimerization Domain at 1.65-Å Resolution 2003).
a. Cette liaison induit une courbure de 59° de l'ADN, facilitant l'organisation de la chromatine centromérique (The centromere comes into focus: from CENP-A nucleosomes to kinetochore connections with the spindle 2020).
(Figure : vetopsy.fr d'après Kixmoeller et coll)
b. CENP-B agit en tant que dimère, ce qui lui permet de lier deux boîtes CENP-B distantes.
Cette capacité de pontage est essentielle à la formation de boucles d'ADN submicroniques entre les répétitions α-satellites, contribuant ainsi à la compaction et à l'organisation tridimensionnelle de la chromatine centromérique (CENP-B-mediated DNA loops regulate activity and stability of human centromeres 2022).
Rôles de CENP-B
Bien que CENP-B ne soit pas indispensable à la formation d'un centromère fonctionnel, elle favorise l'assemblage de novo et contribue au maintien des centromères contenant de l'ADN α-satellite (CENP-B-mediated DNA loops regulate activity and stability of human centromeres 2022).
1. Cette absence de caractère indispensable est illustrée par deux situations.
a. Les " néocentromères " sont des centromères fonctionnels qui apparaissent exceptionnellement sur une région chromosomique initialement dépourvue d'activité centromérique, le plus souvent à la suite d'une réorganisation chromosomique ou de l'inactivation d'un centromère préexistant.
Ils se forment généralement en l'absence d'ADN α-satellite et de boîtes CENP-B, tout en recrutant CENP-A, le CCAN et un kinétochore complet, assurant ainsi une ségrégation correcte des chromosomes.
b. De même, le centromère du chromosome Y humain contient peu ou pas de boîtes CENP-B fonctionnelles.
Malgré cette particularité, il recrute normalement CENP-A et les autres constituants du kinétochore, ce qui confirme que la présence de CENP-B constitue un facteur favorisant, mais non une condition nécessaire à la formation d'un centromère fonctionnel.
Ces deux exemples montrent que l'identité du centromère est principalement déterminée par la chromatine à CENP-A et les mécanismes épigénétiques qui la maintiennent.
(Figure : vetopsy.fr)
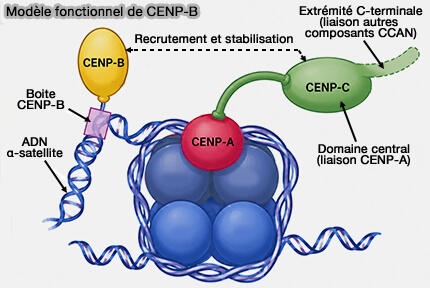
2. Pour favoriser l'assemblage et le maintien du centromère, CENP-B constitue un lien fonctionnel entre l'ADN α-satellite et la chromatine centromérique en interagissant avec :
- CENP-A, dont elle favorise le maintien d'un état de chromatine centromérique ouvert, propice au recrutement des facteurs centromériques (CENP-A and CENP-B collaborate to create an open centromeric chromatin state 2023),
- CENP-C, contribuant à l'organisation du CCAN et à l'assemblage du kinétochore (DNA sequence-specific binding of CENP-B enhances the fidelity of human centromere function 2015).
(Figure : vetopsy.fr d'après Nagpal et coll)
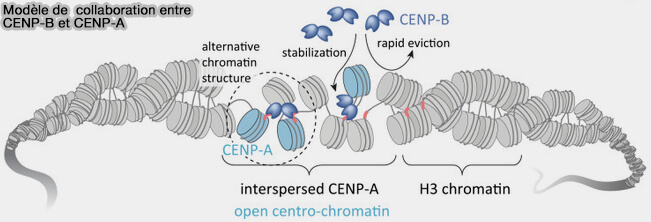
2. Des travaux récents ont montré que les boîtes CENP-B ne sont pas réparties de manière homogène au sein du centromère (Conservation of dichromatin organization along regional centromeres 2025).
a. La plupart des boîtes CENP-B sont méthylées sur la cytosine ( méthylation de l'ADN).
b. En revanche, dans la région de creux du centromère (Centromere Dip Region, CDR), plusieurs boîtes CENP-B adjacentes demeurent hypométhylées, un état associé à une chromatine plus ouverte.
Les dimères de CENP-B s'y fixent préférentiellement et interagissent avec CENP-A et CENP-C, contribuant à l'organisation de la chromatine centromérique et à l'assemblage du kinétochore.
(Figure : vetopsy.fr d'après Fritz Parls)
3. Un modèle bipartite du centromère a été proposé (Vertebrate centromeres in mitosis are functionally bipartite structures stabilized by cohesin 2024).
Un kinétochore humain fixe simultanément 15 à 20 microtubules, qui exercent des forces de traction importantes et répétées.
- Les auteurs proposent qu'une organisation en un seul domaine centromérique ne permettrait pas de répartir efficacement ces contraintes mécaniques.
- Ils suggèrent donc que le centromère est constitué de deux sous-domaines maintenus rapprochés par les cohésines, permettant de répartir ces forces et de limiter les déformations de la chromatine centromérique.
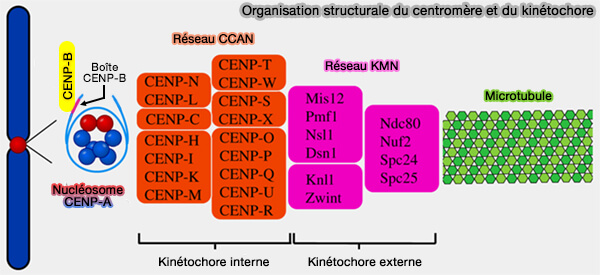
Vers le réseau CCAN
La reconnaissance de l'ADN α-satellite par CENP-B et l'organisation de la chromatine centromérique constituent une étape préparatoire à l'assemblage du réseau constitutif du centromère (CCAN ou Constitutive Centromere-Associated Network).
Ancré au nucléosome contenant CENP-A, le CCAN forme la plateforme du kinétochore interne et recrute le réseau KMN (KNL1–Mis12–Ndc80), qui assure l'attachement des microtubules au kinétochore.
BiochimieChimie organiqueBioénergétiqueAcides nucléiquesNucléotidesADNARNChromatineStructure dynamique de la chromatineNucléosomesHistonesMembres des histonesCode des histonesEuchromatine, hétérochromatine et remodelageChromosomesStructure des chromosomesCentromèreTélomèreProtidesGlucidesLipidesEnzymesCoenzymesVitaminesHormonesComposés inorganiquesTransport membranaireMoteurs moléculairesVoies de signalisation