EHD2 (Eps15-Homology Domain containing protein 2) est une protéine EHD, ATPase liée à la dynamine située au col des cavéoles, dont la fonction physiologique majeure est la stabilisation cavéolaire à la membrane.
Remarque : des protéines transmembranaires comme ROR1 (Receptor tyrosine kinase-like Orphan Receptor 1), qui interagit avec CAV1, et la cavine 1 pourraient faciliter la formation de cavéoles, mais pas dans tous les tissus.
2. La composition lipidique de nanodomaines membranaires, en particulier, cholestérol et en phospholipides chargés négativement, i.e. phosphatidylsérine (PS) et PI(4,5)P2 ou PIP2, est indispensable à la biogenèse des cavéoles.
L'assemblage ainsi que les interactions entre toutes ces molécules, i.e. protéines/lipides, protéines/protéines et lipides/lipides sont loin d'être connus, ce qui implique que certains articles un peu anciens ne sont plus d'actualité et que les modèles sont nombreux.
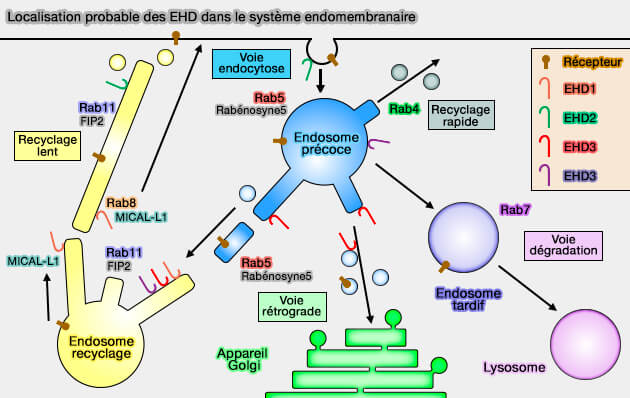
Localisation probable des EHD dans le système endomembranaire
(Figure : vetopsy.fr d'après Naslavsky et coll)
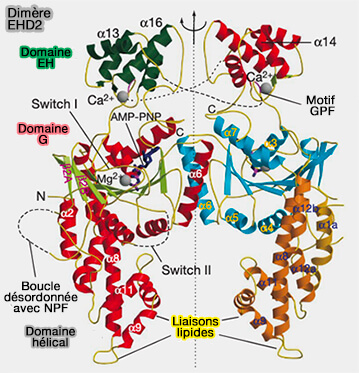
Structure de EHD2
EHD2 (Eps15-Homology Domain containing protein 2) est une protéine EHD, ATPase liée à la dynamine située au col des cavéoles, qui comprend plusieurs domaines.
2. Un domaine charnière est représenté par Pro286 (Pro289 dans EHD4) qui pivote lors de l'activation (Hinge) pour exposer les sites de liaison membranaire, i.e. α9 et α11.
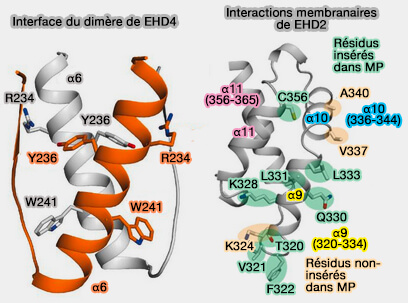
3. Le domaine hélicoïdal, i.e. α8-α12 (287-398), contient les sites de liaison membranaire.
Les dimères EHD2 interagissent avec les membranes via des interactions ioniques médiées par α9-α11 sur une interface incurvée.
La courbure de membrane imposée, perpendiculaire à la courbure imposée par la face de liaison de membrane concave du dimère EHD2, pourrait entraîner un stress de courbure dans la bicouche lipidique et faciliterait ainsi le réarrangement lipidique nécessaire à la formation d'étapes intermédiaires vers la fission/fusion membranaire.
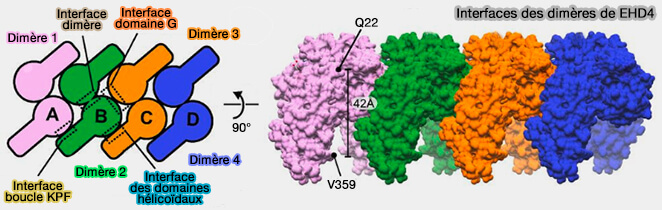
Interactions de EHD4
(Figure : vetopsy.fr d'après Melo et coll)
Les changements conformationnels induits par l'hydrolyse des nucléotides conduisent à une déstabilisation supplémentaire de la membrane.
4. Le lien (400-446) contient le motif GPF qui est impliqué dans l'auto-inhibition du dimère.
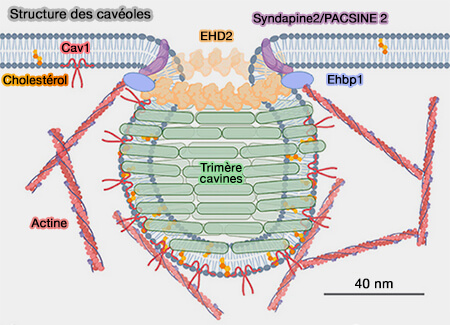
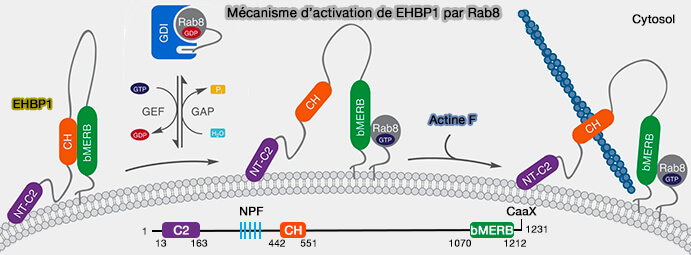
Ehbp1 cavéolaire et actine
(Figure : vetopsy.fr d'après Matthaeus et coll)la limitation de la mobilité des cavéoles à l'intérieur de la membrane.
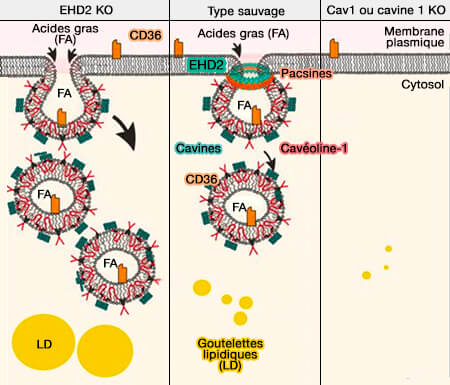
1. EHD2, comme les autres EHD, joue un rôle dans la formation des cavéoles en régulant négativement ce processus de bourgeonnement favorisé par le cholestérol ( cavéoline et cholestérol).
De faibles niveaux d'expression d'EHD2 sont liés à l'obésité.
2. La perte d'EHD2 conduit à la scission des cavéoles, vraisemblablement par hydrolyse de l'ATP qui désassemble les oligomères d'EDH2, ce qui permet le recrutement de la dynamine ( détachement des cavéoles).
dans le remodelage des membranes plasmiques lors de l'augmentation de tension par des changements de courbure de la membrane et le renforcement des fibres de stress.
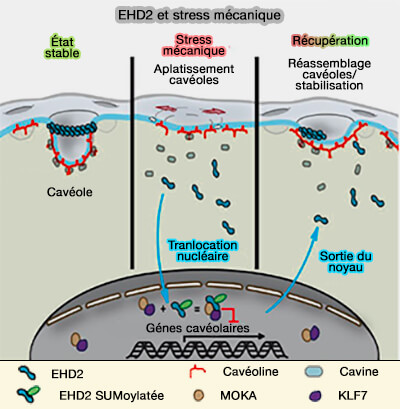
Lors de stress mécanique, Cav1, la cavine 1 et EHD2 sont libérés des cavéoles aplaties.
La seule EHD2 est SUMOylée et subit une translocation vers le noyau où elle régule la transcription des gènes par l'interaction avec MOKA et KLF-7.
Lorsque le stress mécanique s'achève, EHD2 sort du noyau et est nécessaire à la stabilisation du réservoir des cavéoles au niveau de la membrane plasmique.
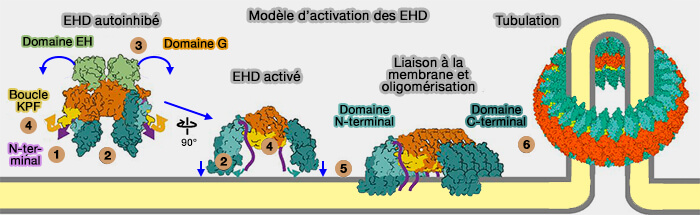
Activation de EHD2
1. En solution, EHD2 se dimérise en conformation inhibée, en particulier grâce au motif GPF ( inhibition des EHD).
2. Lorsque EHD2 est recruté sur les membranes par sa liaison à l'ATP, une série de changements conformationnels aligne les sites de liaison des phospholipides avec la membrane et facilite l'oligomérisation de l'EHD2 en structures annulaires (activation des EHD où le mécanisme est détaillé).
Modèle d'activation des EHD
(Figure : vetopsy.fr d'après Melo et coll)
3. Lors de l'hydrolyse de l'ATP, l'oligomère EHD se désassemble et se déplace de la membrane des cavéoles vers le cytosol.




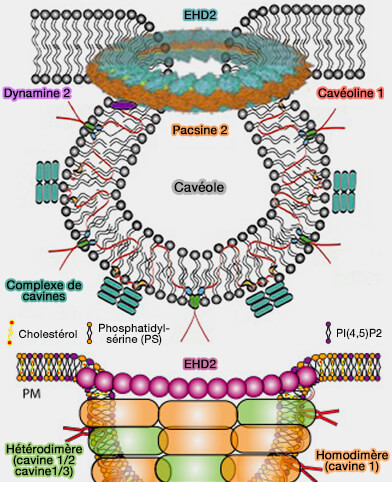

 oligomérisation des EHD).
oligomérisation des EHD).