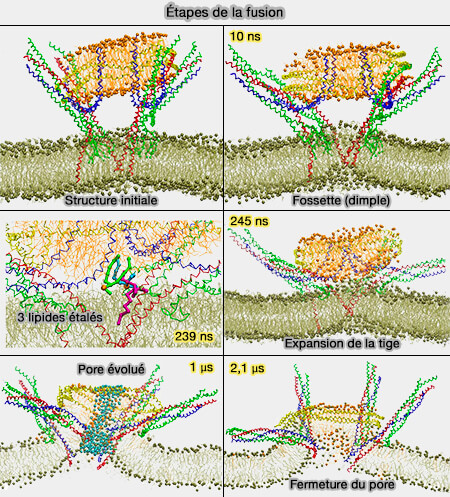
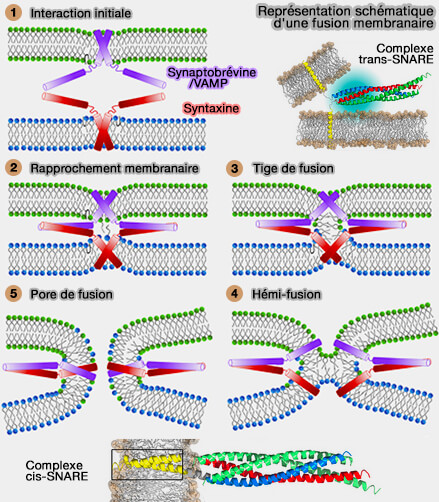
Suite à la formation de la tige de fusion, i.e. aboutissant à la fusion des feuillets externes membranaires, les feuillets internes des membranes doivent fusionner pour former le pore de fusion.
La fusion des vésicules nécessite une machinerie de fusion conservée par l'évolution et composée de plusieurs protéines ( machinerie de fusion).
Pore de fusion initial
La fusion complète des vésicules nécessite que les feuillets internes fusionnent pour former un pore de fusion initial qui se trouve dans un état d'énergie minimale locale, i.e. il est métastable.
Les pores sont généralement considérés comme des structures lipidiques symétriques en forme de sablier bordées de protéines de fusion, ce qui est loin d'être le cas ( asymétrie des pores).
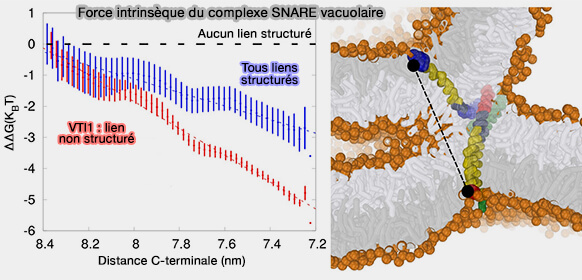
Le complexe SNARE des vacuoles de levure transmet approximativement une force de 18 pN aux feuillets internes d'environ 8 nm d'épaisseur de la tige de fusion, lorsque les trois SNARE contenant les TMD, i.e. Nyv1, R-SNARE, Vam3 et Vti1, Q-SNARE, l'autre Q-SNARE étant Vam7, sont dans une structure α-hélicoïdale.
L'amplitude de cette force est réduite de moitié à 9 pN lorsque le lien de Vti1 adopte une structure non hélicoïdale.
La compréhension des rôles et des mécanismes des protéines de fusion est intimement liée à l'énergie libre des intermédiaires formés lors de la réaction de fusion, i.e. c'est un des problèmes centraux à résoudre à l'heure actuelle.
Deux modèles de formation du pore de fusion
Les TMD des SNARE subissent des forces de traction et/ou de flexion et les transmettent à la tige de fusion, ce qui permet la formation des pores de fusion selon deux modèles.
L'extrémité C-terminale hydrophile des TDM des SNARE est tirée dans le noyau hydrophobe de la ou des membranes.
L'ajout d'agents hydrophiles ou de résidus chargés entrave ce processus.
2. Le modèle par indentation suppose que les extrémités C-terminales favorisent la progression de la fusion en exerçant des forces ponctuelles sur la membrane, facilitant des déformations fortes et hautement localisées des feuillets internes.
Force intrinsèque du complexe SNARE vacuolaire
(Figure : vetopsy.fr d'après D'Agostino et coll)
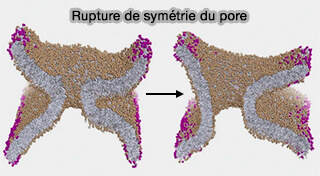
Asymétrie des pores
Le pore de fusion est généralement supposé être axialement symétrique et toroïdal.
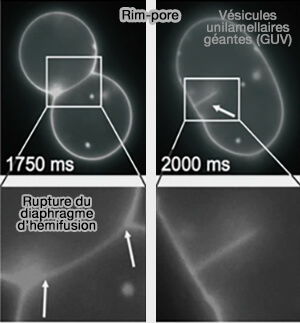
1. Les rim-pore facilitent l'oscillation des pores (pore flickering).
Les extrémités des TMD des SNARE exercent une force directe sur le bord du HD, ce qui favorise la formation d'un pore et la rupture de symétrie concomitante.
Seule une partie du pore est constituée par le bord libre de la membrane avec un coût énergétique élevé.
L'autre partie implique la membrane continue énergétiquement moins coûteuse, i.e. le col d'un pore de fusion toroïdal.
En raison de sa nature hétérogène, la forme d'un tel pore n'est pas circulaire et sa forme générale lors de l'expansion est une conséquence directe de la minimisation de l'énergie libre.
Cette barrière peut être assez faible, si bien que des fluctuations thermiques peuvent faciliter des fluctuations substantielles de la taille et de la conductance des pores.
Une telle étape d'oscillation des pores est suivie d'une expansion complète et soudaine si l'expansion des pores dépasse la taille critique des pores.
Formation d’un rim-pore au bord (rim) d’un diaphragme d’hémifusion
(Figure : vetopsy.fr d'après Risselada et coll)
3. Les rim-pores peuvent se fermer en l'absence d'une barrière d'énergie libre, en particulier en l'absence de tension de la membrane.
Par conséquent, les événements dits " kiss-and-run ", i.e. l'ouverture et la fermeture transitoires des pores de fusion, dans les systèmes de vésicules synaptiques pourraient également s'expliquer par la présence de pores de bordure dans les diaphragmes d'hémifusion de petite à moyenne taille, i.e. de diamètre de 10 nm ou plus, plutôt que par des pores de fusion toroïdaux.
En particulier, il est difficile d'imaginer comment la fermeture d'un pore de fusion toroïdal via des événements d'hémifission pourrait se produire en l'absence d'une machinerie de fission pilotée par GTP.
4. La formation et la fermeture des pores du bord pourraient entraîner une conductance transitoire des ions.
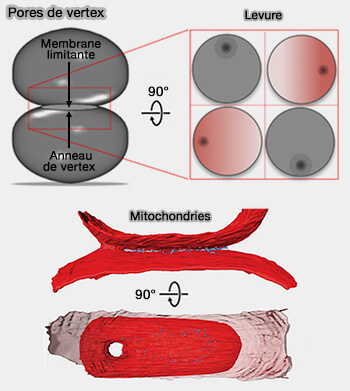
Pores de vertex
(Figure : vetopsy.fr d'après D'agostino et Brandt)
Les comportements de " kiss-and-run " seraient stimulés par les mêmes facteurs qui améliorent la survie et la croissance d'un petit diaphragme d'hémifusion, tels qu'un nombre réduit de complexes SNARE participants au site de fusion membranaire et la présence d'une concentration élevée de cholestérol.
5. In vivo, la formation de pores de fusion se déroule de manière compétitive entre les pores de bord et les pores de fusion toroïdaux, i.e. l'équilibre entre ces deux voies est plutôt soumis à des conditions externes telles que la composition de la membrane et les niveaux d'expression des protéines.
En outre, ce pore de fusion reliant au centre deux feuillets incurvés membranaires brisera favorablement sa symétrie et s'agrandira pour atteindre son équilibre.
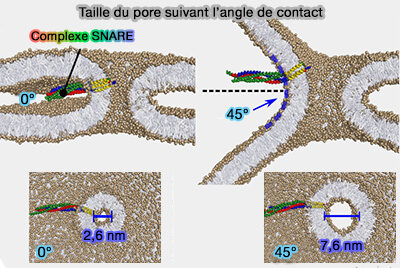
2. La taille adoptée par le pore de vertex résultant dépend fortement de l'angle de contact apparent entre les vésicules en contact, même en l'absence de tension superficielle de la membrane.
Taille du pore suivant l’angle de contact
(Figure : vetopsy.fr d'après Blokhuis et coll)
Les grands angles de contact accroissent l'équilibre du pore.
Parce que la machinerie de fusion des membranes cellulaires ancre activement les membranes, elle facilite une expansion collective de la zone de contact et augmente l'angle de contact.





 machinerie de fusion).
machinerie de fusion).