Le pyruvate, $\ce{CH3COCOO-}$, est une molécule située au carrefour de plusieurs voies métaboliques dont la perturbation joue un rôle essentiel dans de nombreuses affections, en particulier les cancers par l’effet Warburg dans lequel le pyruvate est transformé en lactate.
Le pyruvate, $\ce{CH3COCOO-}$, est une molécule située au carrefour de plusieurs voies métaboliques dont la perturbation joue un rôle essentiel dans de nombreuses affections :
Pyruvate
(Figure : vetopsy.fr)1. les cancers par l'effet Warburg,
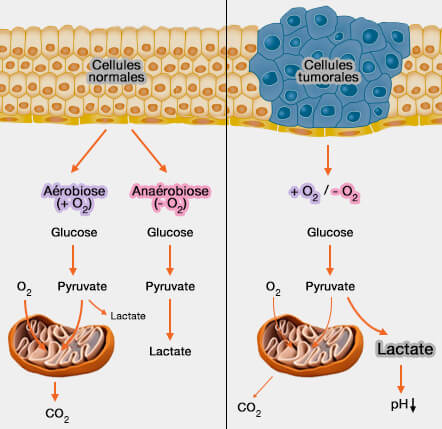
La LDH (lactose déshydrogénase) joue un rôle important dans l'effet Warburg, du nom du médecin, physiologiste et biochimiste allemand Otto Heinrich Warburg (1880-1970), prix Nobel de physiologie ou médecine en 1931, dont un de ses étudiants fut un certain Hans Adolf Krebs (1900-1981), prix Nobel en 1953.
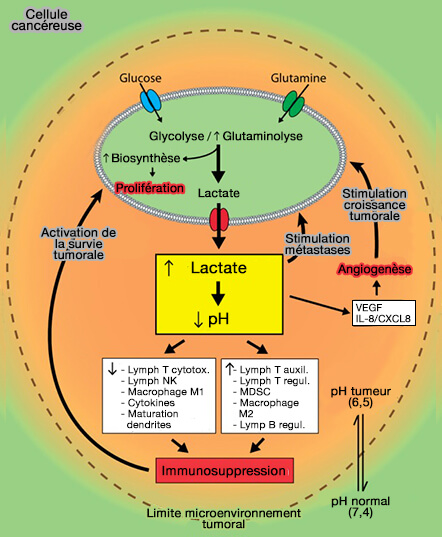
b. L'exportation de lactate hors de la cellule, facilitée par les transporteurs de monocarboxylate (MCT1 et MCT4), symports lactate/$\ce{H+}$, acidifie le milieu entourant les cellules cancéreuses.
c. Le lactate peut être utilisé comme source de combustible par les cellules cancéreuses situées à la surface de la tumeur, où, après conversion en pyruvate, les niveaux d'oxygène sont suffisants pour supporter la phosphorylation oxydative.
Elle favorise la synthèse d'acides aminés et des nucléotides nécessaires à la prolifération cellulaire pour shunter la glycolyse.
Le $\ce{CO2}$ libéré conduit à l'acidification de la matrice extracellulaire par sa combinaison avec l'eau ($\ce{HO2}$) pour produire des protons ($\ce{H+}$) et du bicarbonate ($\ce{HCO3-}$) grâce aux anhydrases carboniques, et en particulier, à l'anhydrase carbonique 9 (CA9/CAIX) associée à la membrane. Les protons sont aussi exportés grâce à l'antiport $\ce{Na+}$/$\ce{H+}$ 1 (NHE-1 ou SLC9A1).
les fibroblastes qui activent les métalloprotéases et la hyaluronidase pour dégrader la matrice extracellulaire et créer un microenvironnement tumoral favorable à la migration des cellules tumorales (métastases).
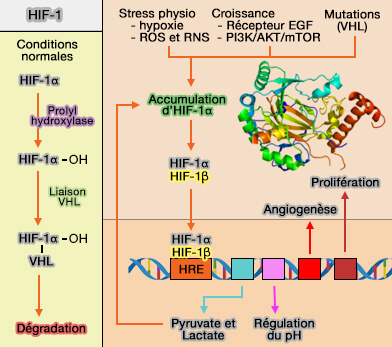
2. Dans les cellules cancéreuses où p53 demeure silencieux, le processus conduit à l'élévation de la glycolyse, i.e. la formation de pyruvate et de lactate.
3. Au-delà de son rôle dans la régulation métabolique, p53 est également un régulateur central du cycle cellulaire, des checkpoints, de l’apoptose et de la sénescence, notamment par l’induction de gènes effecteurs comme p21 ( p53 et activation de p21).
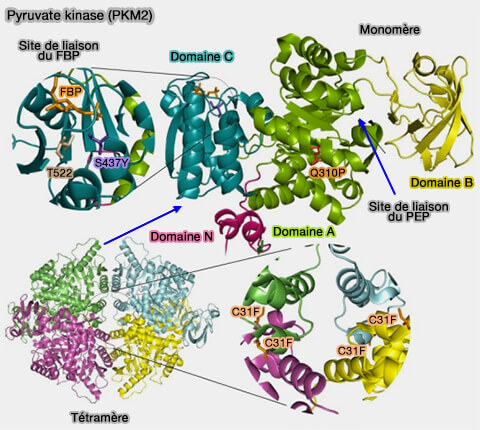
Les cellules cancéreuses modifient directement le métabolisme du pyruvate en déplaçant le profil d'expression des enzymes glycolytiques, en particulier celui de la pyruvate kinase (PK).
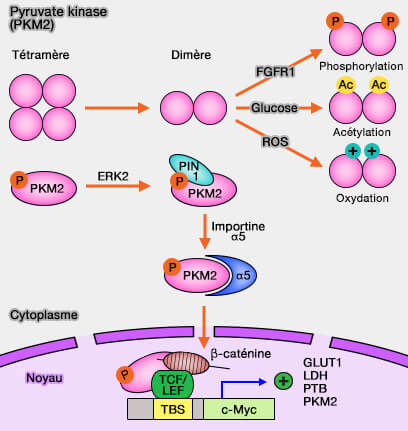
PKM2 et régulation
(Figure : vetopsy.fr d'après Yang et coll)
Toutefois, PKM2 est la seule à pouvoir former des dimères qui sont inactifs : l'interconversion entre dimères et tétramères est modulée par diverses modifications post-traductionnelles, par des effecteurs allostériques et des partenaires de liaison (Regulation and function of pyruvate kinase M2 in cancer 2013).
PKM2, sous forme dimérique, redirige efficacement le flux de carbone de la production de pyruvate et de l'énergie cellulaire vers les voies anabolisantes nécessaires à la croissance cellulaire rapide (PKM2, a Central Point of Regulation in Cancer Metabolism 2013).
Les cellules cancéreuses favorisent la formation de PKM2 dimérique (i.e. inactive) qui provoque l'accumulation d'intermédiaires de la glycolyse. Ces intermédiaires sont introduits dans d'autres voies, et en particulier celle des pentoses phosphates.
Elle produit des équivalents réducteurs sous la forme de NADPH, qui peuvent être utilisés pour soutenir la biosynthèse des lipides et la régénération du glutathion-H2, i.e glutathion réduit.
Le glutathion réduit est utilisé pour diminuer le stress oxydatif des cellules cancéreuses, qui est supérieur à celui des cellules normales, pour éviter leur mort cellulaire.






 MCT1 et MCT4), symports lactate/$\ce{H+}$, acidifie le milieu entourant les cellules cancéreuses.
MCT1 et MCT4), symports lactate/$\ce{H+}$, acidifie le milieu entourant les cellules cancéreuses.