


Glucides : dégradation du glucose
Glycolyse ou voie d'Embden-Meyerhof
Phase préparatoire
- Biochimie
- Chimie organique
- Bioénergétique
- Composition de la matière vivante
- Transport membranaire
- Moteurs moléculaires
- Voies de signalisation

La production d'énergie à partir du glucose est essentiellement assurée par la voie d'Embden-Meyerhof.

Cette voie doit son nom au biologiste et chimiste allemand Gustav Embden (1874-1933) et au physicien et biochimiste allemand Otto Fritz Meyerhoff (1884-1951), qui seul fut prix Nobel de physiologie et de médecine en 1922 ( Histoire de la glycolyse).
Histoire de la glycolyse).
On y rajoute souvent le nom du biologiste soviétique Jakub Karol Parnas (1884-1945).
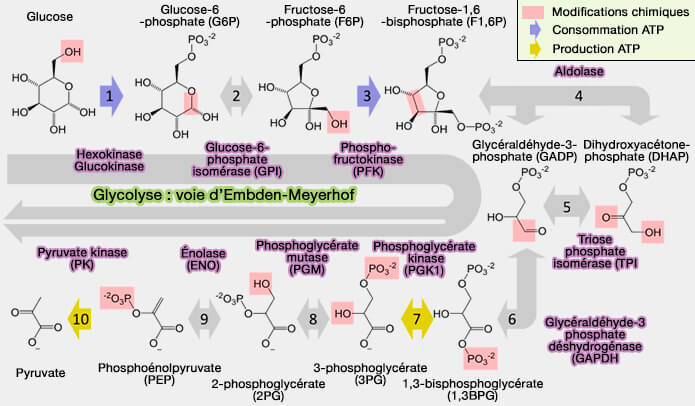
Vue d'ensemble de la glycolyse
1. La voie d'Embden-Meyerhof-Parnas conduit au pyruvate, molécule commune à tous les organismes :
$\ce{Glucose + 2 NAD+ + 2 ADP + 2 Pi}$ $\longrightarrow$ $\ce{2 pyruvate + 2 NADH + 2 H+ + 2 ATP + 2 H2O}$
L'avenir du pyruvate dépendra des cellules selon qu'elles fonctionnent (devenir du pyruvate) :
- en aérobiose, i.e. avec de $\ce{O2}$, i.e. en présence d'une chaîne respiratoire de transport d'électrons, le pyruvate est oxydé en acétyl-CoA (décarboxylation oxydative dans la matrice mitochondriale) pour entrer dans le cycle de Krebs (cycle de l'acide citrique,
- en anaérobiose, i.e. sans $\ce{O2}$, le pyruvate est réduit au cours de fermentations conduisant essentiellement au lactate (fermentation lactique) et à l'éthanol (fermentation alcoolique).
(Figure : vetopsy.fr d'après Thomas Shafee)
{kind=link}
2. La glycolyse, qui se déroule entièrement dans le cytosol, est divisée en deux phases principales :
- une phase dite préparatoire qui conduit du glucose à deux glycéraldéhyde-3-phosphate (G3P), avec consommation d'ATP,
- une phase finale, oxydo-réduction couplée à la formation du pyruvate et d'ATP.
Toutes les réactions sont, en principe, réversibles et équilibrées, exceptées les trois réactions catalysées par des kinases qui sont fortement énergétiques et nécessitent d'autres systèmes enzymatiques pour être inversées :
- l'hexokinase ou la glucokinase, qui phosphoryle le glucose en glucose-6-phosphate (réaction 1) ;
- la phosphofructokinase, qui phosphoryle le fructose-6-phosphate en fructose-1,6-bisphosphate (réaction 3) ;
- la pyruvate kinase (PK), qui transfère le groupe phosphate du phosphoénolpyruvate (PEP) à l'ADP pour former de l'ATP et du pyruvate (réaction 10).
3. D'autres oses que le glucose peuvent entrer dans le cycle d'Embden-Meyerhof ( autres glucides de la glycolyse), à savoir :
- des polysaccharides de réserve comme le glycogène par la glycogénolyse (ou l'amidon chez les plantes),
- d'autres oses comme le mannose, le fructose ou le galactose.
Phase préparatoire de la glycolyse
Cette phase conduit à 2 molécules de glycéraldéhyde-3-phosphate à partir d'une molécule de glucose.
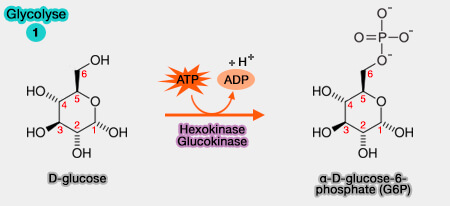
1. Phosphorylation du glucose en glucose-6-phosphate
Le glucose, après pénétration dans le cytosol, est phosphorylé en position 6 pour obtenir du glucose-6-phosphate.
$\ce{Glucose + ATP ->[Mg++] glucose-6-phosphate + ADP + H+}$
(Figure : vetopsy.fr)
Cette réaction, dont le co-facteur est l'ion $\ce{Mg++}$, nécessite la présence d'une kinase.
1. L'hexokinase (EC 2.7.1.1) intervient dans la plupart des tissus, en particulier dans les muscles des mammifères.
- Elle est peu spécifique car elle phosphoryle aussi le mannose, le fructose ou la glucosamine.
- Les isoformes de l'hexokinase (HKI, HKII et HKIII) ont une forte affinité pour le glucose (KM inférieure à 1 mM).

L'hexokinase est inhibée par le glucose-6-phosphate, son effecteur, ce qui constitue une régulation essentielle de la glycolyse (régulation de la glycolyse).
Les interactions hexokinase/VDAC, les VDAC (Voltage Dependent Anion Channel) étant des porines, canaux de la membrane externe mitochondriale (OMM), lient la glycolyse à la phosphorylation oxydative. La liaison de HK1 au VDAC augmente l'efficacité catalytique des deux processus :
- en facilitant la libération d'ATP mitochondrial par les VDAC pour la phosphorylation du glucose,
- en canalisant l'ADP dans les mitochondries pour la phosphorylation oxydative (The catalytic inactivation of the N-half of human hexokinase 2 and structural and biochemical characterization of its mitochondrial conformation 2018).
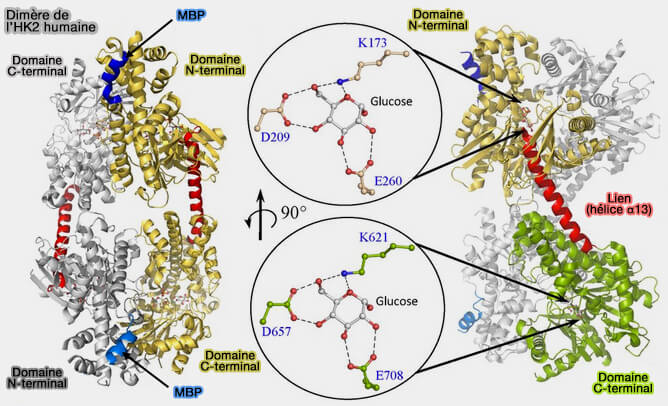
(Figure : vetopsy.fr d'après Nawaz et coll)
2. La glucokinase (EC 2.7.1.2), étroitement spécifique du glucose, est essentiellement hépatique, mais aussi pancréatique, et intervient de façon efficace que lorsque le taux de glucose sanguin est élevé, par exemple après un repas (régulation de la glycolyse).
- Son affinité pour le glucose est beaucoup plus faible que celle de l'hexokinase (KM égale à 10 mM).
- Sa mutation intervient dans certains diabètes (50% des diabètes MODY - maturity-onset diabetes of the young -).
a. Du point de vue hépatique,
- lors d'hyperglycémie, le foie utilise la glucokinase pour transformer le glucose en glucose-6-P pour la glycogénogenèse (stockage du glucose sous forme de glycogène),
- lors d'hypoglycémie, le glucose sera métabolisé par d'autres organes grâce à l'héxokinase car la glucokinase est inactive.
b. Du point de vue pancréatique, l'hyperglycémie provoque la transformation du glucose en glucose-6-P par la glucokinase, ce qui induira la sécrétion d'insuline (hormone hypoglycémiante).
3. La réaction de phosphorylation du glucose est fortement exergonique ($\ce{\Delta G'0}$ d'environ -4 kcal.mole-1).
- La réaction est donc irréversible dans les conditions physiologiques.
- La formation de glucose à partir du glucose-6-phosphate nécessite l'intervention de la glucose-6-phosphatase (EC 3.1.3.9).
Le glucose-6-phosphate est le produit de départ dans la voie des pentoses phosphates, l'une des quatre voies métaboliques principales du métabolisme énergétique.
4. Le glucose-6-phosphate reste prisonnier de la cellule, car il ne possède pas de transporteur spécifique. Il peut également provenir :
- de la transformation du glucose-1-phosphate libéré lors de la glycogénolyse ( étape 2 de la glycogénolyse),
- lors d'interconversions à partir d'autres oses, grâce à des transcétolases ou des transaldolases pour arriver au glycéraldéhyde-3-phosphate (voie des pentoses et photosynthèse).
Remarque : le glucose-6-phosphate peut aussi être converti en glucose-1-phosphate (G1P) pour être converti en glycogène.
2. Isomérisation du glucose-6-phosphate
en fructose-6-phosphate
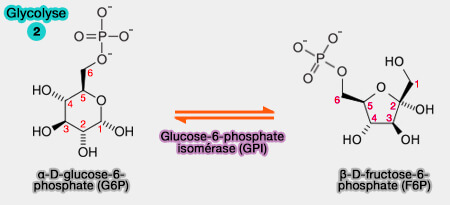
(Figure : vetopsy.fr)
1. L'Isomérisation d l'α-D-glucose-6-phosphate en β-D-fructose-6-phosphate est réversible et catalysée par la glucose-6-phosphate isomérase (GPI) ou phosphohexose isomérase (EC 5.3.1.9).
$\ce{Glucose-6-phosphate}$ $\leftrightharpoons$ $\ce{Fructose-6-phosphate}$
L'équilibre est en faveur de la formation de glucose-6-phosphate car le fructose-6-phosphate est rapidement transformé par la réaction suivante.
2. La réaction inverse est effective dans la voie des pentoses phosphates.
3. Formation de fructose-1,6-bisphosphate
Le β-D-fructose-6-phosphate est phosphorylé en β-D-fructose-1,6-bisphosphate (F1,6BP).
$\ce{Fructose-6-phosphate} + ATP$ $\longrightarrow$ $\ce{fructose-1,6-biphosphate + ADP}$
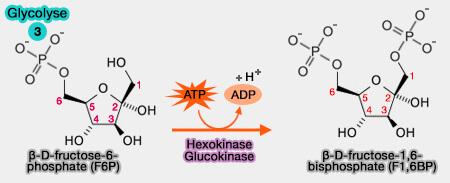
(Figure : vetopsy.fr)
1. Cette réaction, dont le co-facteur est l'ion $\ce{Mg++}$, comme la réaction de phosphorylation du glucose nécessite la présence de la phosphofructokinase 1 (PFK-1), i.e. EC 2.7.1.11.
PFK-1 fait partie des phosphofructokinases, qui comprennent aussi la phosphofructokinase 2 (PFK-2).
La réaction est fortement exergonique ($\ce{\Delta G'0}$ d'environ -4 kcal.mole-1) : c'est la réaction la plus lente de la glycolyse.
- La réaction est donc irréversible dans les conditions physiologiques.
- La formation de fructose-6-phosphate à partir du fructose-1,6-phosphate nécessite l'intervention de la fructose-1,6-bisphosphatase (régulation de la glycolyse).
2. La phosphofructokinase est une enzyme allostérique tétramérique qui contient pour chaque protomère :
- deux sites actifs, un pour le fructose-6-phosphate et 1 pour l'ATP,
- un site de contrôle allostérique en fonction de divers effecteurs dont l'ATP.
Elle joue un rôle important dans le contrôle de la glycolyse (régulation de la glycolyse).
4. Formation des trioses phosphates
1. La coupure du β-D-fructose-1,6-bisphosphate conduit à deux molécules de trioses phosphates : le D-glycéraldéhyde-3-phosphate (GADP) et la dihydroxyacétone phosphate (DHAP).
$\ce{β-D-fructose-1,6 bisphosphate}$ $\leftrightharpoons$ $\ce{D-glycéraldéhyde-3-phosphate + dihydroxyacétone phosphate}$
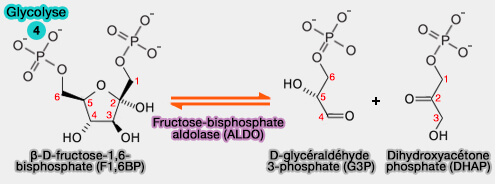
(Figure : vetopsy.fr)
Cette réaction est catalysée par la fructose-bisphosphate aldolase ou aldolase B (EC 4.1.2.13).
- La réaction est réversible, ce qui permet la synthèse des hexoses à partir des trioses phosphates.
- Au cours de la réaction, au moins chez les animaux et les plantes (aldolase de classe 1), il se forme une base de Schiff ($\ce{R2C=NR'}$) entre le groupe carbonyle ($\ce{C=O}$) du dihydroxyacétone phosphate et un groupe ε-aminé d'une lysine de la protéine enzymatique : le groupe ε-aminé ($\ce{NH3+}$) est attaché au quatrième carbone, compté à partir du carbone α ou 1.
Remarque : Cette enzyme intervient à la fois dans :
- la glycolyse,
- la gluconéogenèse,
- le cycle de Calvin de la photosynthèse.
2. L'-somérisation du glucose-6-phosphate en fructose-6-phosphate est réversible et catalysée par la glucose-6-phosphate isomérase (GPI) ou phosphohexose isomérase (EC 5.3.1.).
Remarque : DHAP peut être converti en glycérol-3-phosphate :
- soit pour produire du glycérol,
- soit pour entrer dans la synthèse des triglycérides et des glycérophospholipides ( synthèse des triglycérides et des glycérophospholipides).
5. Interconversion des trioses phosphates
Les deux trioses phosphates, le D-glycéraldéhyde-3-phosphate (GADP) et le dihydroxyacétone phosphate (DHAP), sont interconvertibles.
$\ce{dihydroxyacétone phosphate$ $\leftrightharpoons$ $\ce{D-glycéraldéhyde-3-phosphate}$
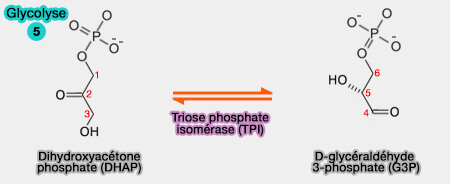
(Figure : vetopsy.fr)
Cette réaction est catalysée par une triose phosphate isomérase (TPI ou TIM), EC 5.3.1.1.
- L'équilibre de la réaction est en faveur de la formation de dihydroxyacétone phosphate.
- Mais, comme le G3P est transformé rapidement par la réaction suivante de la glycolyse, l'équilibre est constamment déplacé et l'isomérase peut alors assurer la transformation de la totalité du dihydroxyacétone phosphate.
De ce fait, à la fin de cette phase préparatoire, deux molécules de glycéraldéhyde-3-phosphate sont formées à partir d'une molécule de glucose.