La perturbation du pyruvate dans les astrocytes explique une grande partie de la pathogénie des maladies neurodégénératives, et en particulier de la maladie d'Alzheimer.
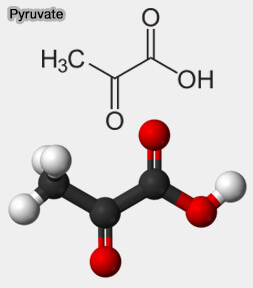
Le pyruvate, $\ce{CH3COCOO-}$, est une molécule située au carrefour de plusieurs voies métaboliques dont la perturbation joue un rôle essentiel dans de nombreuses affections :
Pyruvate
(Figure : vetopsy.fr)1. les cancers par l'effet Warburg,
1. L'apport d'énergie au cerveau dépend presque entièrement du métabolisme du glucose et donc, du pyruvate : son fonctionnement requiert 20-25 % de la consommation quotidienne de glucose du corps.
La perturbation de toutes les fonctions des astrocytes dans les maladies neurodégénératives explique les symptômes observés (maladies neurodégénératives).
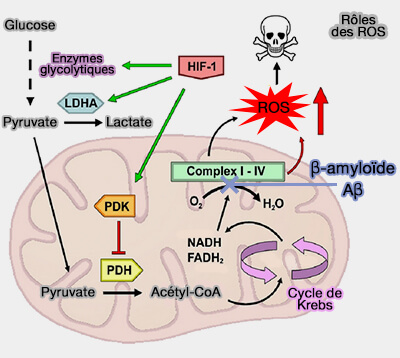
La production des dérivés réactifs de l'oxygène (ROS) est considérée comme l'une des caractéristiques de la pathogénie de la maladie d'Alzheimer.
Ces ROS jouent un rôle important dans la signalisation cellulaire et l'homéostasie, mais leur accumulation entraîne des dommages cellulaires importants, d'où le nom de stress oxydatif.
Le résultat final de la production des ROS neuronaux est une diminution de l'activité de la pyruvate déshydrogénase (PDH), diminuant drastiquement la production d'ATP et provoquant un dysfonctionnement neuronal essentiel à la pathogénie de la maladie d'Alzheimer.
Lorsque l'absorption du glucose est perturbée, les astrocytes manquent d'énergie pour éliminer suffisamment le glutamate de la fente synaptique.
Ce processus entraîne une neurotoxicité excitatrice qui provoque, entre autres, l'augmentation des ROS et un dysfonctionnement mitochondrial provoquant la mort neuronale (Astrocytes and Epilepsy 2016).
Ces régions comprennent le cortex préfrontal bilatéral, le lobe pariétal latéral bilatéral, le cingulum/précunéum postérieur, le gyrus rectum, le gyrus temporal latéral bilatéral et les noyaux caudés bilatéraux.
En revanche, le cervelet et le gyrus temporal inférieur ont des niveaux significativement plus bas de glycolyse.
Réseau du mode par défaut (Default Mode Network ou DMN)
(Figure : vetopsy.fr d'après Andreashorn)
1. Le DMN réseau est mobilisé lorsque l'individu n'est pas engagé dans une tâche précise, comme lors de réveries où l'esprit vagabonde : elle consomme la majorité de l'énergie cérébrale et synchronise l'ensemble des aires cérébrales qui sont prêtes à exécuter une tâche.
Ce réseau fut découvert par hasard par l'imagerie cérébrale sur des patients à qui l'on demandait de ne penser à rien pour avoir un référentiel lors des examens.
Lors d'une activité particulière, le cerveau ne mobilise que 5 % d'énergie de plus que cette activité de " repos ".
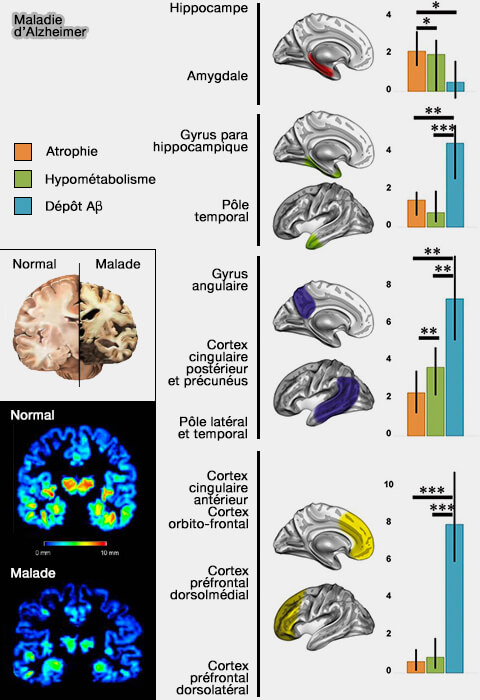
L'absence de corrélation significative entre l'atrophie et l'hypométabolisme peut indiquer le fonctionnement de mécanismes pathologiques ou protecteurs spécifiques à une région.
En conclusion, l'augmentation de la glycolyse aérobie pourrait compenser le dysfonctionnement neuronal et l'hypométabolisme.





 fonctions des astrocytes).
fonctions des astrocytes).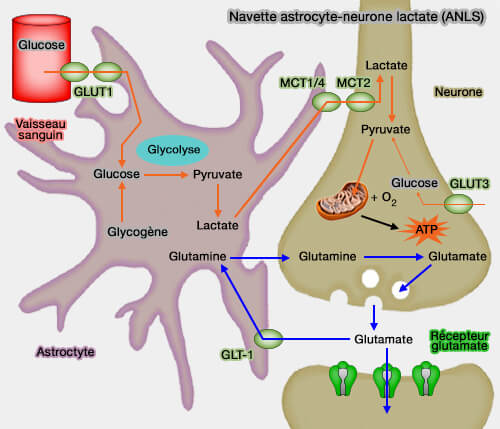



{kind=link}