Les protéines CENP-E, CENP-F et CENP-J assurent des fonctions spécialisées au cours de la division cellulaire, en dehors du réseau constitutif du centromère (CCAN).
Certaines interviennent de manière transitoire au niveau du kinétochore pendant la mitose, tandis que d'autres remplissent des fonctions distinctes au niveau du centrosome.
CENP-F participe à l'organisation du kinétochore et à la progression de la mitose.
CENP-J (CPAP) est principalement une protéine du centrosome essentielle à la biogenèse des centrioles.
CENP-E
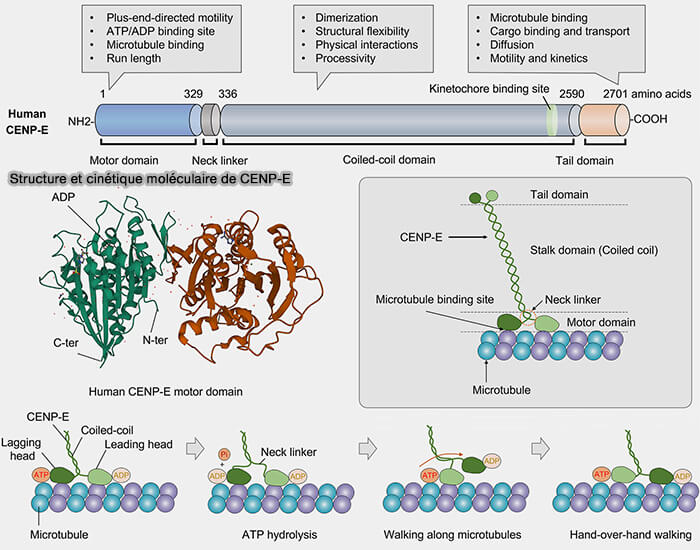
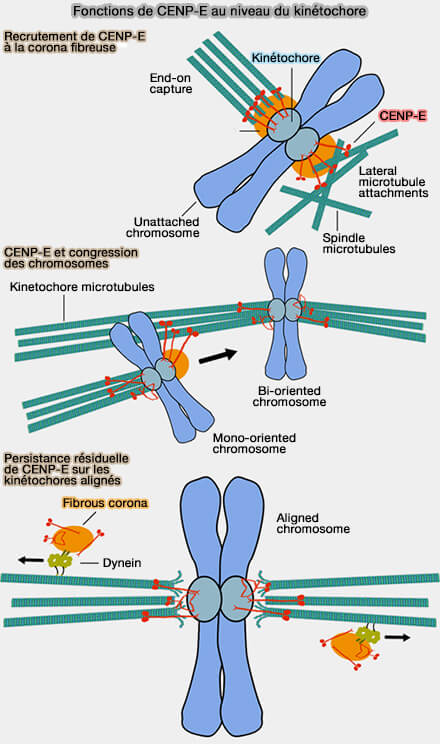
CENP-E (Centromere Protein E)/KIF10 est une kinésine mitotique de grande taille appartenant à la famille des kinésines-7, localisée au kinétochore externe exclusivement au cours de la mitose, où elle est essentielle à la capture, à l'attachement et à l'alignement des chromosomes sur la plaque équatoriale.
Son activité ATPase est stimulée par l'interaction avec les microtubules.
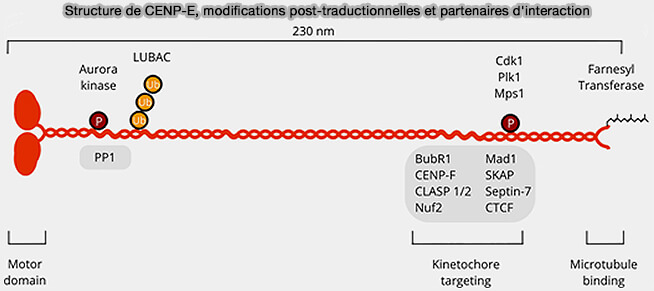
2. Le domaine central coiled-coil (340-2100) forme une longue tige flexible assurant la dimérisation de CENP-E, la transmission des forces entre le domaine moteur et le domaine C-terminal, ainsi que sa régulation conformationnelle.
CENP-E adopte une conformation repliée (" folded ") inactive en interphase,
Elle se déploie en une conformation étendue (" extended ") active en mitose.
Activités motrices dans l'assemblage du fuseau
(Figure : vetopsy.fr d'après Yang et coll)
3. Le domaine C-terminal (résidus 2100–2600) assure le recrutement et l'ancrage de CENP-E au kinétochore externe.
a. Il interagit avec plusieurs protéines de la corona, notamment :
Tant que tous les chromosomes ne sont pas correctement attachés et soumis à une tension suffisante, CENP-E contribue au maintien du signal inhibiteur qui retarde le déclenchement de l'anaphase.
Régulation de CENP-E
1. L'activité et la localisation de CENP-E sont régulées par plusieurs phosphorylations mitotiques, notamment par CDK1, Aurora B et Mps1, qui modulent son recrutement, son activité motrice et ses interactions avec le kinétochore.
Structure de CENP-E, modifications post-traductionnelles et partenaires d'interaction
(Figure : vetopsy.fr d'après Craske et Welburn)
Plusieurs inhibiteurs spécifiques de CENP-E sont actuellement étudiés comme agents anticancéreux, notamment GSK923295, le seul à avoir atteint un essai clinique de phase I, ainsi que Syntelin et PF-2771, encore au stade préclinique.
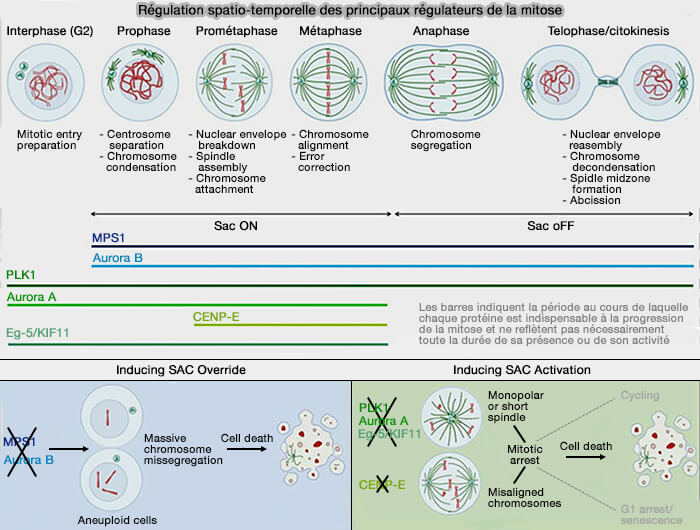
Régulation spatio-temporelle des principaux régulateurs de la mitose
(Figure : vetopsy.fr d'après Novais et coll)
CENP-F
CENP-F (Centromere Protein F) est une protéine centromérique transitoire, localisée au kinétochore externe durant la mitose.
Contrairement à CENP-E, qui est une protéine motrice assurant le déplacement des chromosomes, CENP-F agit principalement comme une protéine d'échafaudage contribuant à l'organisation de la corona fibreuse, à la stabilisation des attachements entre les kinétochores et les microtubules et au bon déroulement de la ségrégation des chromosomes.
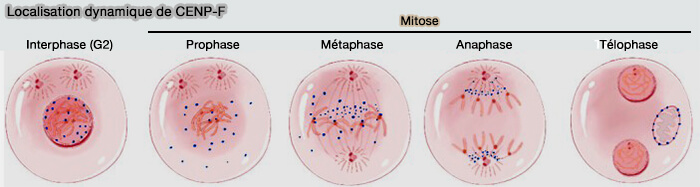
Localisation dynamique de CENP-F
(Figure : vetopsy.fr d'après Wan et coll)
Structure de CENP-F
CENP-F est une très grande protéine d'environ 3 100 à 3 200 résidus selon les isoformes, constituée principalement des domaines coiled-coil lui conférant une structure filamentaire flexible qui lui permet d’interagir avec plusieurs partenaires protéiques.
1. Le domaine N-terminal assure le recrutement de CENP-F au kinétochore grâce à des interactions avec plusieurs protéines recrutées au kinétochore au cours de la mitose, notamment :
2. Le domaine central, riche en régions coiled-coil, assure la dimérisation de CENP-F et forme une longue tige flexible favorisant les interactions avec de nombreux partenaires protéiques.
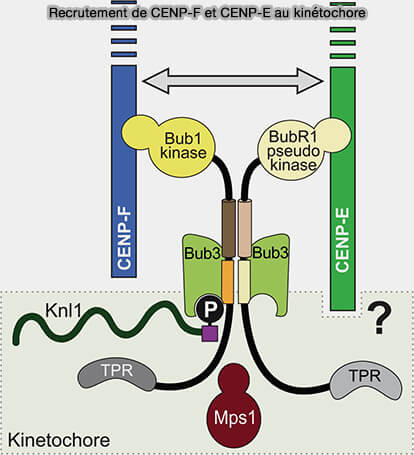
Recrutement de CENP-F et CENP-E au kinétochore
(Figure : vetopsy.fr d'après Ciossani et coll) le KT-core, un domaine essentiel au recrutement de CENP-F au kinétochore, contenant deux leucine zippers, dont l'une renferme une cystéine hautement conservée (C2865 chez l'Homme), indispensable à la liaison au kinétochore,
un signal de localisation nucléaire (NLS), qui favorise l'import de CENP-F dans le noyau et son accumulation à l'enveloppe nucléaire avant la rupture de l'enveloppe nucléaire (NEBD), facilitant ainsi son recrutement ultérieur au kinétochore
un motif CAAX de farnésylation, dont la cystéine est modifiée par l'ajout covalent d'un groupe farnésyle (C15), modification lipidique favorisant le recrutement et la stabilité de CENP-F au niveau du kinétochore au cours de la mitose, sans être indispensable à sa localisation.
En son absence, des défauts d'alignement des chromosomes et une désorganisation de la corona fibreuse sont observés.
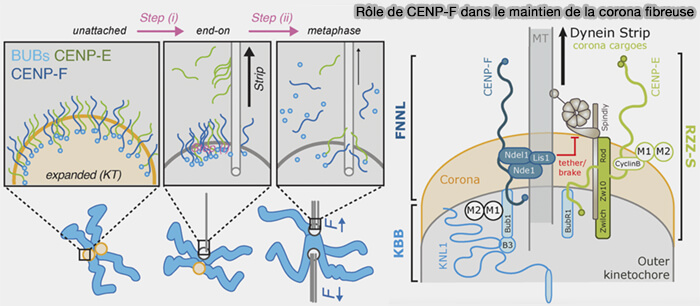
b. Elle participe au maintien de la corona fibreuse en limitant le retrait prématuré (stripping) par la dynéine de plusieurs composants de cette structure, notamment CENP-E et d'autres protéines de la corona fibreuse, contribuant ainsi au bon fonctionnement du kinétochore.
Rôle de CENP-F dans le maintien de la corona fibreuse
(Figure : vetopsy.fr d'après Auckland et coll)
c. Elle coopère avec CENP-E, la dynéine et plusieurs protéines du kinétochore externe afin de coordonner les mouvements chromosomiques et la maturation des attachements entre les kinétochores et les microtubules ( congression et attachements des chromosomes).
L'activité et la localisation de CENP-F sont régulées par plusieurs phosphorylations mitotiques, notamment par CDK1 et Plk1, qui contrôlent son recrutement au kinétochore ainsi que ses interactions avec les autres protéines du kinétochore externe.
1. Une surexpression de CENP-F est observée dans de nombreux cancers, où elle est associée à une instabilité chromosomique accrue, à un potentiel métastatique plus élevé et à un mauvais pronostic (Centromere Protein F in Tumor Biology: Cancer's Achilles Heel 2025).
En raison de son rôle essentiel dans la progression de la mitose, CENP-F constitue également une cible thérapeutique potentielle pour le développement de traitements anticancéreux.
2. Les mutations du gène CENPF sont responsables de plusieurs maladies du développement, notamment du syndrome de Strømme, caractérisé par des anomalies oculaires, une microcéphalie et diverses malformations congénitales.
CENP-J
CENP-J (Centromere Protein J), également appelée CPAP (Centrosomal P4.1-associated Protein), est une protéine localisée principalement au centrosome.
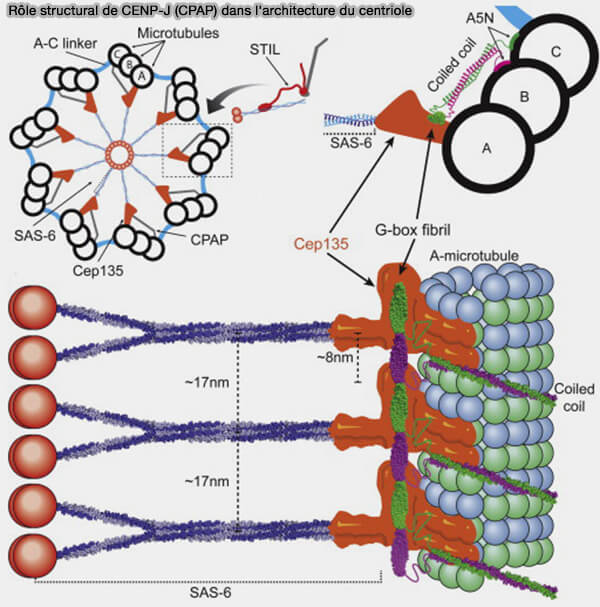
Rôle structural de CENP-J (CPAP) dans l'architecture du centriole
(Figure : vetopsy.fr d'après Hatzopoulos et coll)
Structure de CENP-J
CENP-J est une protéine d'environ 1 330 à 1 340 acides aminés selon les isoformes, constituée principalement de domaines coiled-coil lui conférant une structure allongée favorisant les interactions avec de nombreuses protéines centrosomales.
Le domaine central, riche en régions coiled-coil, assure la dimérisation de CENP-J et sert de plateforme d'interaction avec plusieurs protéines impliquées dans la duplication des centrioles, notamment STIL, CEP120 et CEP135.
Le domaine C-terminal participe au recrutement, au maintien et à la maturation des centrioles, contribuant ainsi à leur stabilité.
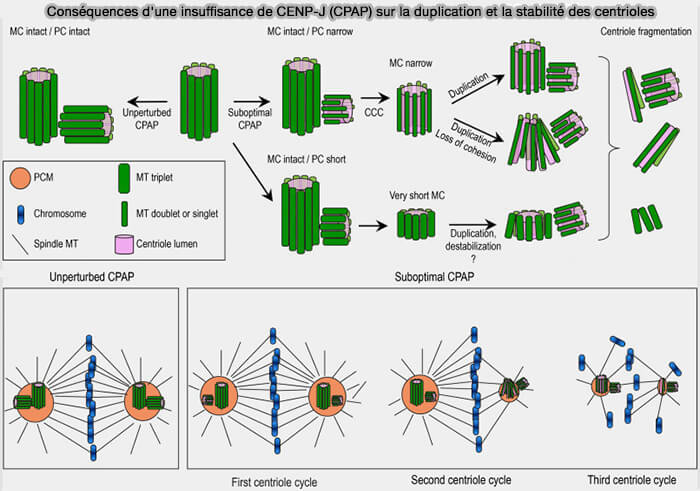
Conséquences d'une insuffisance de CENP-J (CPAP) sur la duplication et la stabilité des centrioles
(Figure : vetopsy.fr d'après Vásquez-Limeta et coll)
b. Elle contrôle ensuite l'élongation des centrioles en régulant la polymérisation des microtubules, contribuant ainsi à leur longueur et à leur stabilité.
Lorsque la quantité ou l'activité de CENP-J est insuffisante, les procentrioles restent plus courts ou incomplets.
Ces anomalies, souvent discrètes lors du premier cycle de duplication, s'amplifient progressivement au cours des cycles cellulaires suivants, entraînant une perte de cohésion des centrioles, leur fragmentation et des défauts croissants d'organisation du fuseau mitotique.
Cette activité est notamment régulée par des phosphorylations dépendantes de PLK4, qui coordonnent la duplication et l'élongation des centrioles au cours du cycle cellulaire.
Les protéines CENP-A à CENP-U constituent un ensemble fonctionnel assurant l'identité du centromère, l'organisation du kinétochore et les interactions avec les microtubules du fuseau.
Le tableau récapitulatif présente leur localisation, leurs principales fonctions et leur dynamique au cours du cycle cellulaire.





 congression et attachements des chromosomes).
congression et attachements des chromosomes).