Les spermatozoïdes sont des cellules sexuelles (ou gamètes) mâles produites, chez les mammifères, par les testicules.
Vous pouvez étudier :
la spermatogenèse qui, à partir d'une spermatocyte
I (diploïde) aboutit à quatre spermatides (haploïdes), grâce à une division particulière
appelée méiose qui réduit le nombre de chromosomes de moitié (n) ;
la spermiogenèse qui transforme les spermatides
en spermatozoïdes ;
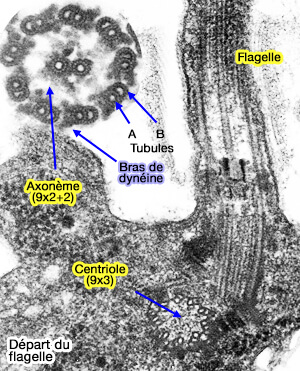
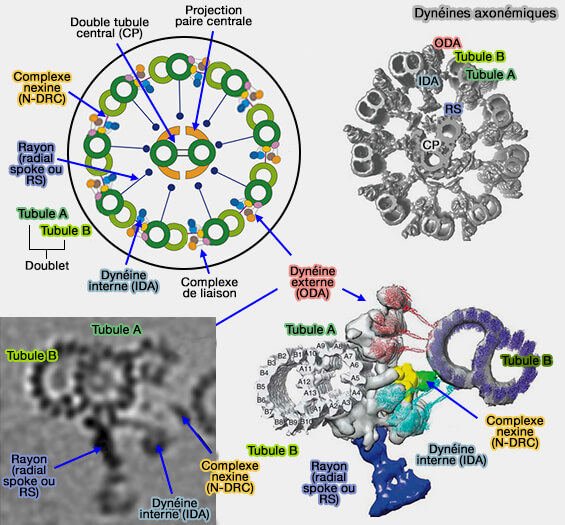
L'axonème forme la superstructure des microtubules du flagelle (ou du cil mobile) et comprend une structure dite " 9+2 " hautement conservée :
9 doublets de microtubules périphériques, i.e. tubule A et B,
2 singulets de microtubules, appelé appareil central ou paire centrale (CP) , de faible longueur, qui se termine dans la partie proximale de l'axonème, entouré d'une gaine centrale.
Les mouvements du flagelle sont dus aux glissements entre les doublets des microtubules périphériques (et non à leurs raccourcissements) les uns par rapport aux autres, grâce à la dynéine.
Doublets et dynéine
1. Les doublets sont composés d'un microtubule cylindrique complet (tubule A de 13 protofilaments) et d'un tubule incomplet (tubule B de 15 protofilaments) attaché au tubule A.
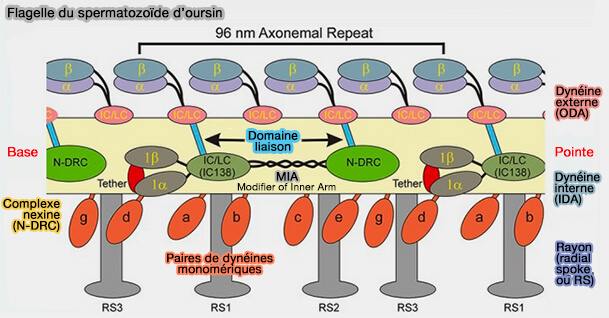
Les doublets adjacents sont liés, i.e. avec une périodicité régulière de 96 nm, par :
des bras de dynéines externes, i.e. ODA ou Outer Dynein Arm, 4 par 24 nm,
des bras de dynéine internes , i.e. IDA ou Inner Dynein Arm
une nexine ou complexe régulateur de dynéine (DRC).
Un rayon radial (radial spoke), fibres à disposition radiaire se terminant par un renflement, est localisé entre le doublet et la paire centrale.
Flagelle du spermatozoïde d'oursin
(Figure : vetopsy.fr d'après King)
2. La dynéine est une énorme protéine qui fait partie des moteurs moléculaires et qui utilise l'ATP comme combustible.
Le glissement du tubule A déplace les doublets voisins.
Le mouvement de flexion de la tige (ou axonème) est également provoqué par les liaisons de nexine.
Si on détruit les ponts de nexine, l'axonème s'allonge (de neuf fois sa longueur) au lieu de se courber.
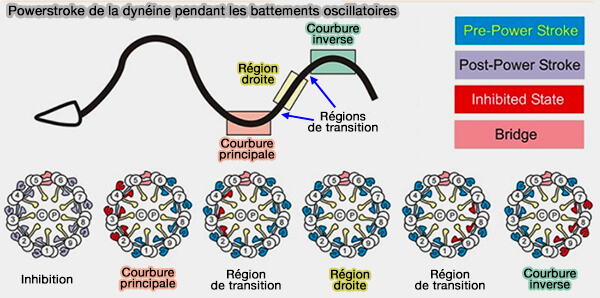
2. La motilité flagellaire, mais aussi ciliaire, est causée par l'action d'ensemble des bras intérieur et extérieur de la dynéine (Turning dyneins off bends cilia 2018)
Les dynéines d'un seul côté de l'axonème fonctionne activement pendant la flexion et cette région motrice propage le mouvement sur toute la longueur axonémale.
La génération de la courbure inversée résulte de la commutation du côté opposé de l'axonème.
Powerstroke de la dynéine pendant les battements oscillatoires
(Figure : vetopsy.fr d'après King)
Les mécanismes encore mal compris sont complexes et expliqués dans :
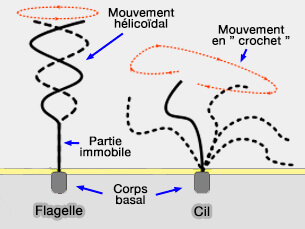
4. Les mouvements des flagelles sont ondulatoires et génèrent des ondes pseudo-hélicoïdales : le spermatozoïde se déplace comme s'il possédait une hélice.
L'onde se déplace le long du flagelle de la tête à la queue.
Les cils, qui possèdent une structure identique, ont des mouvements complexes en 3D, dits " en crochet ", ce qui permet de balayer les liquides ou les particules dans une seule direction, comme dans de nombreux organes du corps, et, en particulier, dans les voies spermatiques.
Une fois dans l'épididyme, le spermatozoïde acquiert sa mobilité.
Dans la tête, la motilité est quasi nulle (le mouvement est vibratile).
Dans le corps, elle progresse au fur à mesure.
Dans la queue de l'épididyme où le spermatozoïde acquiert la mobilité dite fléchante (mouvement linéaire).
Mouvements d'un flagelle et d'un cil
(Figure : vetopsy.fr d'après Kohidai L.)
D'une part, le pH et le Ca++ intracellulaire diminuent lors du parcours, la concentration en AMP cyclique augmente et les fibres denses de la pièce intermédiaire se rigidifient. D'autre part, la modification de la composition du liquide épididymaire (volume fortement diminué, pression osmotique forte au niveau de la tête) participe à la mobilité qui se modifie au cours du trajet des spermatozoïdes. En outre, la température de l'épididyme (-7° par rapport au corps chez le rat) conduit également à une diminution de cette mobilité.
A la fin de l'épididyme, des spermatozoïdes peuvent se grouper, tous dans le même sens, en faisceau d'une dizaine pour augmenter leur vitesse.
Leur vitesse est multipliée par trois environ (140 µm/s), ce qui leur permet d'atteindre l'ovule plus rapidement.
Mouvements dans le système génital femelle
Les mouvements des spermatozoïdes augmentent dans la cavité utérine où ils subissent la capacitation (cf. fécondation)
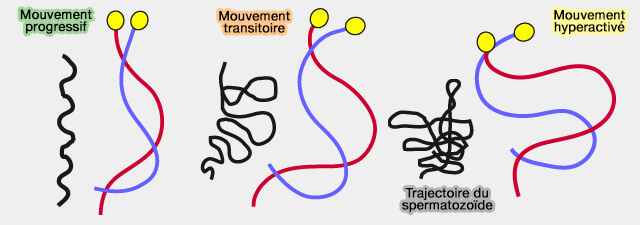
Ce mouvement hyperactivé consiste en une augmentation de le fréquence et de l'amplitude des battements flagellaires : le mouvement devient très rapide et de large amplitude (trajectoire typique en forme de créneau).
Cette hyperactivité est due à la perte de cholestérol, l'augmentation de l'entrée des ions Ca++, la phosphorylation des protéines flagellaires.
Différents mouvements des spermatozoïdes
(Figure : vetopsy.fr)
Les mouvements des spermatozoïdes deviennent " frénétiques " et leur permettent :
de se détacher s'ils sont collés sur la muqueuse de l'utérus pour attendre le moment de l'ovulation (comme chez la chienne),






 mouvements de la dynéine).
mouvements de la dynéine).