


Acides nucléiques
ARN : traduction
Élongation, terminaison et régulation
- Biochimie
- Chimie organique
- Bioénergétique
- Composition de la matière vivante
- Transport membranaire
- Moteurs moléculaires
- Voies de signalisation

L'élongation, la terminaison et la régulation de la traduction assurent la synthèse, l'arrêt et le contrôle de la production des protéines ainsi que le recyclage des ribosomes.
Après l'assemblage du ribosome 80S au cours de l'initiation de la traduction, plusieurs mécanismes successifs assurent la synthèse, l'arrêt et le contrôle de la production des protéines.
Ces mécanismes comprennent :
- l'élongation de la traduction,
- la terminaison de la traduction,
- le recyclage des ribosomes,
- les mécanismes de surveillance et de contrôle qualité de la traduction,
- les mécanismes de régulation de la traduction.
Élongation de la traduction
L'élongation correspond à la phase de synthèse de la chaîne polypeptidique au cours de laquelle les acides aminés sont incorporés successivement selon l'ordre des codons présents dans l'ARN messager (ARNm)
À l'issue de l'initiation, le ribosome 80S est assemblé, l'ARN de transfert (ARNt) initiateur chargé de méthionine (Met-ARNtiMet) est positionné dans le site P et le site A est libre pour accueillir le premier ARNt aminoacylé ( séquence CCA à l'extrémité 3' de l'ARNt).
séquence CCA à l'extrémité 3' de l'ARNt).
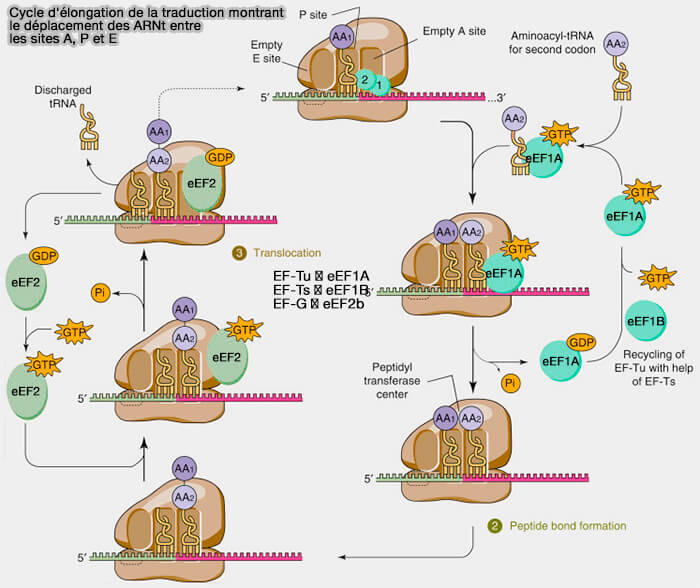
1. Chaque cycle d'élongation débute par l'arrivée dans le site A d'un ARNt aminoacylé transporté par le facteur d'élongation eEF1A associé au GTP (Functions and Regulation of Translation Elongation Factors 2022).
a. eEF1A assure le recrutement des ARNt chargés vers les ribosomes et participe au contrôle de la fidélité de la traduction.
- Seuls les ARNt dont l'anticodon établit un appariement correct avec le codon exposé dans le site A du ribosome déclenchent efficacement l'hydrolyse du GTP porté par eEF1A, permettant leur incorporation stable dans le ribosome.
- Les ARNt incorrectement appariés induisent plus difficilement les changements conformationnels nécessaires à cette hydrolyse et se dissocient généralement avant la formation de la liaison peptidique.
b. La reconnaissance correcte du codon déclenche l'hydrolyse du GTP porté par eEF1A et la libération de ce facteur.
eEF1A-GDP est ensuite recyclé par eEF1B, un GEF (Guanine nucleotide exchange factor) qui permet le remplacement du GDP par un nouveau GTP.
(Figure : vetopsy.fr adaptée d'après Molecular Cell Biology 2016 de Doherty et Guo)
2. Après l'incorporation de l'ARNt aminoacylé dans le site A, le centre peptidyl-transférase (PTC) situé à l'interface des sites A et P de la sous-unité 60S catalyse la formation d'une liaison peptidique entre l'acide aminé porté par l'ARNt du site A et la chaîne polypeptidique attachée à l'ARNt du site P.
- Cette réaction transfère la chaîne polypeptidique en croissance sur l'ARNt situé dans le site A, qui devient alors le nouveau peptidyl-ARNt.
- Contrairement à la plupart des réactions biologiques catalysées par des protéines, cette activité est assurée principalement par l'ARNr 28S de la grande sous-unité ribosomique, ce qui fait du ribosome un ribozyme.
3. Après la formation de la liaison peptidique, le ribosome doit se déplacer d'un codon le long de l'ARN messager afin de permettre la poursuite de la traduction (tRNA Translocation by the Eukaryotic 80S Ribosome and the Impact of GTP Hydrolysis 2018).
a. Cette étape, appelée translocation, est assurée par le facteur d'élongation eEF2 associé au GTP (Time-resolved cryo-EM visualizes ribosomal translocation with EF-G and GTP 2021).
L'hydrolyse du GTP provoque des changements conformationnels qui entraînent le déplacement du ribosome dans le sens 5' → 3' de l'ARNm.
b. La translocation s'accompagne du déplacement coordonné des ARNt au sein du ribosome.
- L'ARNt porteur de la chaîne polypeptidique passe du site A vers le site P.
- L'ARNt déchargé passe du site P vers le site E,
- Le site A devient à nouveau disponible pour accueillir un nouveau complexe eEF1A-GTP-ARNt aminoacylé.

Ce cycle de recrutement des ARNt, de formation des liaisons peptidiques et de translocation se répète de manière successive jusqu'à ce qu'un codon stop soit rencontré dans le site A du ribosome.
Terminaison de la traduction
1. La terminaison de la traduction débute lorsqu'un codon stop atteint le site A du ribosome.
- Contrairement aux codons spécifiant des acides aminés, les codons stop UAA, UAG et UGA ne sont reconnus par aucun ARNt.
- Leur présence dans le site A empêche donc le recrutement d'un nouvel ARNt aminoacylé et met fin aux cycles d'élongation.
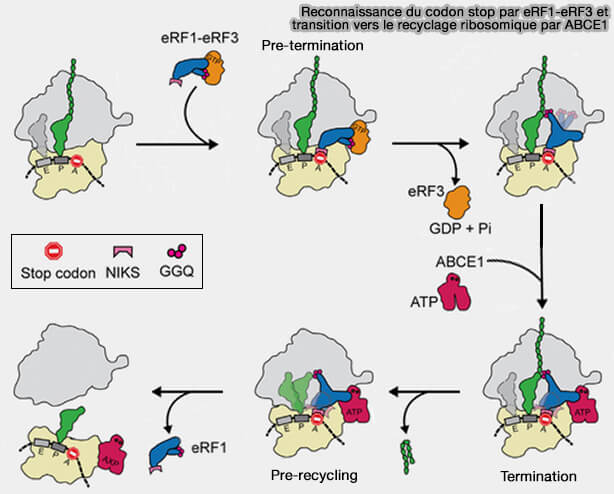
2. Chez les eucaryotes, la reconnaissance des codons stop présents dans le site A, repose sur des interactions directes entre plusieurs motifs conservés du facteur de terminaison eRF1 (eukaryotic Release Factor 1) et le centre de décodage de la sous-unité 40S (Cryoelectron Microscopic Structures of Eukaryotic Translation Termination Complexes Containing eRF1-eRF3 or eRF1-ABCE1 2019).
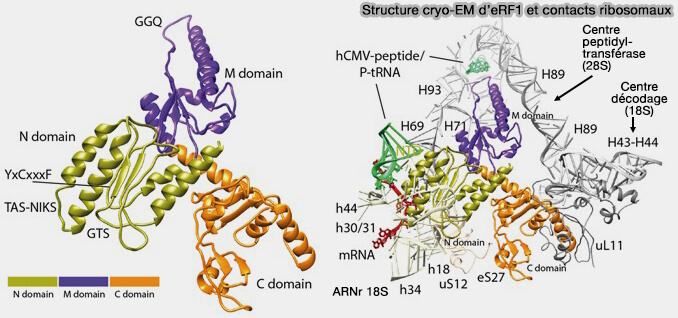
a. Bien que dépourvu d'anticodon, eRF1 adopte une organisation tridimensionnelle mimant celle d'un ARNt, ce qui lui permet d'occuper le site A du ribosome.
- Des motifs conservés de son domaine N, notamment NIKS, GTS et YxCxxxF, établissent des contacts spécifiques avec les trois nucléotides du codon stop et avec l'ARNr du centre de décodage.
- Ces interactions permettent de reconnaître les codons UAA, UAG et UGA tout en excluant les codons codant pour des acides aminés.
b.. La reconnaissance du codon stop est alors transmise à travers la structure de eRF1 jusqu'à son domaine M contenant le motif catalytique GGQ (deux glycines et une glutamine), qui est positionné dans le centre peptidyl-transférase (PTC) de la sous-unité 60S afin de catalyser l'hydrolyse de la liaison ester reliant la chaîne polypeptidique à l'ARNt du site P.
(Figure : vetopsy.fr adaptée d'après Preis et coll)
3. eRF1 est recruté au niveau du site A en association avec eRF3, une GTPase qui facilite le processus de terminaison (The Translation Termination Factor eRF3 Interacts Sequentially with eRF1 and ABCE1 on the Ribosome 2025).
a. La reconnaissance du codon stop par le complexe eRF1-eRF3 déclenche l'hydrolyse du GTP associé à eRF3 et provoque des réarrangements conformationnels qui favorisent l'activation de eRF1 (Structure of a human translation termination complex 2015).
- Cette réorganisation permet au domaine M de eRF1 contenant le motif universel GGQ (Gly-Gly-Gln) de pénétrer dans le centre peptidyl-transférase (PTC) de la sous-unité 60S, notamment au voisinage de l'hélice H89 de l'ARNr 28S, plaçant le facteur de terminaison dans une conformation catalytiquement compétente pour la libération de la chaîne polypeptidique (Conformation of methylated GGQ in the Peptidyl Transferase Center during Translation Termination 2018).
- Le motif NIKS et les régions responsables de la reconnaissance du codon stop demeurent au niveau du centre de décodage de la sous-unité 40S, en interaction avec les hélices h43 et h44 de l'ARNr 18S.
(Figure : vetopsy.fr adaptée d'après Mathheisl et coll)
b. L'hydrolyse de la liaison ester rompt l'attachement covalent entre la chaîne polypeptidique et l'adénosine A76 de l'extrémité 3' CCA de l'ARNt du site P.
- Cette réaction est catalysée au niveau du centre peptidyl-transférase (PTC) de la sous-unité 60S.
- Le motif GGQ de eRF1 confère au centre peptidyl-transférase une activité de peptidyl-ARNt hydrolase en positionnant une molécule d'eau qui devient le nucléophile responsable de l'hydrolyse de la liaison ester.
c. La protéine nouvellement synthétisée est alors libérée du ribosome tandis que l'ARNt devient déacylé car son extrémité 3' CCA n'est plus estérifiée par un acide aminé ou une chaîne peptidique.
4. Le ribosome demeure toutefois associé à l'ARNm ainsi qu'à l'ARNt du site P, formant un complexe post-terminaison.
Ce complexe devra ensuite être démantelé par les mécanismes de recyclage ribosomique.
Recyclage des ribosomes
Après la terminaison de la traduction, les sous-unités ribosomiques, l'ARN messager (ARNm) et les ARN de transfert (ARNt) doivent être dissociés afin de permettre leur réutilisation au cours de nouveaux cycles de synthèse protéique (Ribosome recycling is coordinated by processive events in two asymmetric ATP sites of ABCE1 2018).
Cette étape, appelée recyclage ribosomique, assure le maintien d'un pool fonctionnel de ribosomes disponibles pour l'initiation de nouvelles traductions.
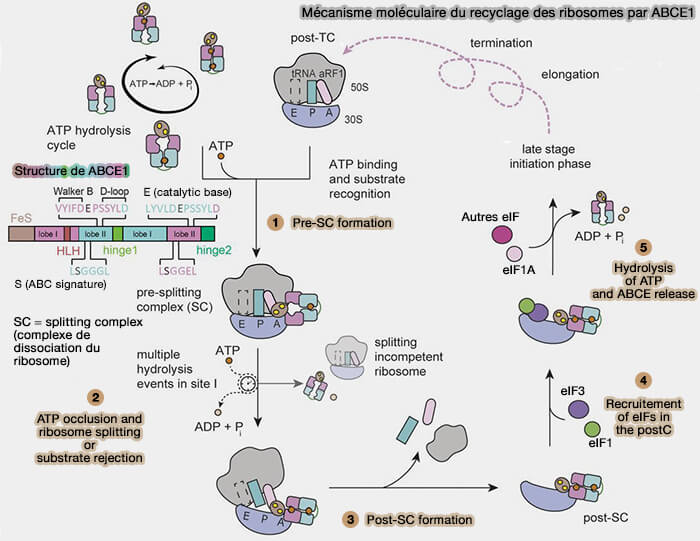
1. Chez les eucaryotes, la protéine ABCE1 joue un rôle central dans ce processus (The Translation Termination Factor eRF3 Interacts Sequentially with eRF1 and ABCE1 on the Ribosome 2025).
a. Cette ATPase de la famille ABC possède un domaine N-terminal contenant deux centres fer-soufre [4Fe-4S] (domaine FeS) ainsi que deux domaines ATPasiques NBD1 et NBD2 (nucleotide-binding domains) formant deux sites de liaison à l'ATP.
- Chaque site ATP est constitué de façon composite par des éléments provenant des deux domaines NBD et comprend notamment un motif signature caractéristique des protéines ABC (LSGGQ), présent chez ABCE1 sous les séquences apparentées LSGGGL et LSGGEL.
- Ces motifs participent à la fermeture des domaines NBD autour des molécules d'ATP et à leur occlusion au sein du complexe.
b. ABCE1 est recrutée sur le complexe post-terminaison en association avec eRF1, qui demeure transitoirement lié au ribosome après la libération de la chaîne polypeptidique.
La fixation et l'hydrolyse de l'ATP par les domaines NBD induisent des changements conformationnels transmis au domaine FeS, ce qui déstabilise les interactions inter-sous-unités et provoque la dissociation du ribosome 80S en sous-unités 40S et 60S distinctes.
(Figure : vetopsy.fr adaptée d'après Nürenberg-Goloub et coll)
2. Après la dissociation des sous-unités, l'ARNm et l'ARNt déacylé demeurent initialement associés à la sous-unité 40S.
Plusieurs facteurs associés à cette sous-unité, notamment eIF1, eIF1A et eIF3, participent alors à leur libération et conduisent à la formation d'une sous-unité 40S recyclée débarrassée des composants résiduels du cycle précédent de traduction.
3. Les sous-unités 40S et 60S rejoignent alors le pool cytoplasmique de ribosomes disponibles pour de nouveaux cycles d'initiation.
Surveillance et contrôle qualité de la traduction
Malgré la grande fidélité des mécanismes de traduction, les ribosomes peuvent rencontrer des ARN messagers (ARNm) présentant des anomalies susceptibles d'interrompre ou de perturber la synthèse protéique.
1. Ces défauts sont généralement détectés au cours de la traduction par des mécanismes capables de surveiller la progression du ribosome, l'intégrité de l'ARNm et le devenir des protéines naissantes, en particulier :
- la présence de codons stop prématurés résultant d'erreurs de transcription, d'épissage ou de mutations génétiques,
- l'absence de codon stop conduisant à la traduction de l'extrémité 3' de l'ARNm, par exemple après une coupure prématurée d'un ARNm ou un défaut de polyadénylation,
- des structures secondaires particulièrement stables ou des lésions de l'ARN capables de bloquer la progression du ribosome, telles que certains pseudonœuds (pseudoknots) des ARN viraux, des tiges-boucles très stables ou des nucléotides endommagés comme les adduits oxydatifs de type 8-oxoguanosine ou les adduits induits par les UV,
- certaines séquences peptidiques ou nucléotidiques provoquant un ralentissement excessif ou un arrêt prolongé de la traduction, notamment des séquences riches en prolines consécutives, certains peptides régulateurs d'arrêt (stalling peptides) ou des régions contenant des codons rares, comme le peptide régulateur humain XBP1u ou des séquences enrichies en codons AGA ou AUA.
2. Les cellules possèdent plusieurs systèmes de surveillance spécialisés capables de détecter ces situations, d'interrompre la traduction, de dissocier les complexes ribosomiques bloqués et d'orienter les ARN messagers ou les protéines aberrantes vers des voies de dégradation adaptées (Mechanisms and functions of ribosome-associated protein quality control 2019).
a. Ces mécanismes comprennent :
- les voies NMD (Nonsense-Mediated Decay), NSD (Non-Stop Decay) et NGD (No-Go Decay), qui assurent la surveillance des ARN messagers,
- les mécanismes de contrôle qualité ribosomique (RQC) qui leur sont associés et assurent la prise en charge des complexes de traduction aberrants, des ribosomes bloqués et des protéines incomplètement synthétisées.
Tous ces mécanismes sont décrits en détail dans la page sur la surveillance des ARN.
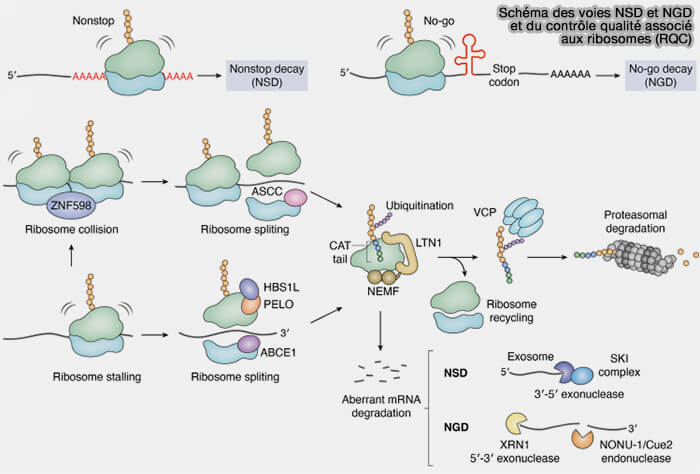
(Figure : vetopsy.fr d'après Ha et coll)
b. L'ensemble de ces mécanismes contribue à préserver la fidélité de l'expression génique, à limiter l'accumulation de protéines aberrantes et à maintenir l'homéostasie protéique de la cellule.
Régulation de la traduction
La traduction n'est pas un processus constitutif mais fait l'objet de nombreux mécanismes de régulation permettant à la cellule d'adapter rapidement la synthèse protéique aux conditions physiologiques et environnementales (Principles of Translational Control: An Overview 2012 et Regulation of Translation Initiation in Eukaryotes: Mechanisms and Biological Targets 2009).
1. La régulation de la traduction intervient principalement au niveau de l'initiation, étape limitante de la synthèse protéique, mais peut également affecter l'élongation, la terminaison, la disponibilité des ribosomes et la stabilité des ARN messagers (The Dynamic Regulation of mRNA Translation and Ribosome Biogenesis During Germ Cell Development and Reproductive Aging 2021).
a. Elle implique de nombreuses voies de signalisation intracellulaires comme :
- la voie mTORC1 (mTORC1, S6K, 4E-BP1), qui stimule la synthèse protéique en réponse à la disponibilité des nutriments, des facteurs de croissance ou de l'énergie cellulaire, (The Translational Regulation in mTOR Pathway 2022),
- les réponses au stress dépendantes de la phosphorylation d'eIF2α (PERK, GCN2, PKR), qui réduisent globalement la traduction lors du stress du réticulum endoplasmique, de la carence en acides aminés, des infections virales ou d'autres conditions cellulaires défavorables (Translation regulation in response to stress 2024).
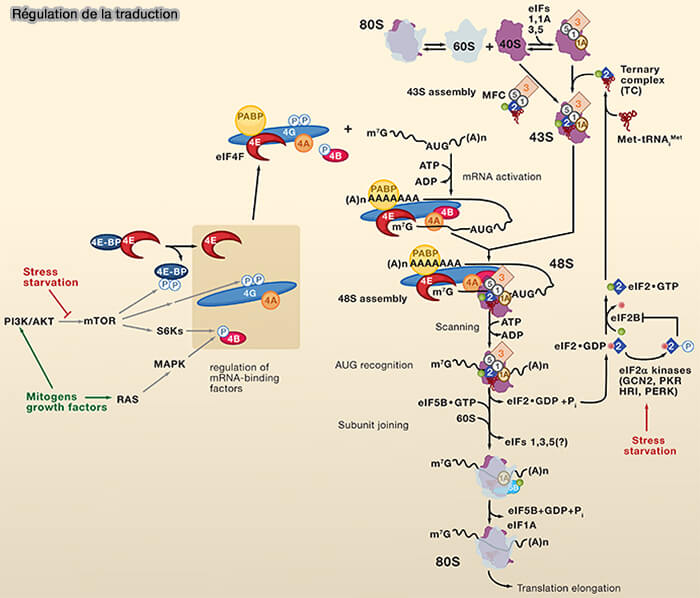
(Figure : vetopsy.fr d'après Sonenberf et Hinnebusch)
b. Elle fait également intervenir :
- des facteurs de traduction (eIF4E, eIF2), dont l'activité contrôle principalement l'initiation de la traduction (Translational regulation by uORFs and start codon selection stringency 2023),
- certaines protéines de liaison aux ARN ou RBP (HuR, TTP), qui modulent notamment la stabilité, la localisation ou la traduction de certains ARN messagers en réponse à différents signaux cellulaires,
- différents ARN régulateurs, notamment les microARN ou miRNA (miR-21, let-7), qui répriment généralement la traduction ou favorisent la dégradation de leurs ARN cibles.
c. Plusieurs modifications chimiques des ARN regroupées sous le terme d'épitranscriptome (m6A, m5C) contribuent également à la régulation de la traduction, de la stabilité et du devenir des ARN (Regulation of translation by site-specific ribosomal RNA methylation 2021).
- Ces modifications sont contrôlées par des protéines writers (METTL3-METTL14), erasers (FTO, ALKBH5) et readers (YTHDF1, YTHDF2), terminologie empruntée au code des histones selon qu'elles assurent respectivement la mise en place, l'élimination ou la reconnaissance des marques chimiques.
- Ces mécanismes participent notamment à l'adaptation de l'expression génique au cours du développement, de la différenciation cellulaire ou des réponses au stress (Regulation of translation by ribosomal RNA pseudouridylation 2023).
2. Ces mécanismes permettent à la cellule d'adapter rapidement la synthèse protéique aux besoins métaboliques, aux signaux extracellulaires, aux conditions de stress ou aux programmes de différenciation.
3. La synthèse d'une chaîne polypeptidique ne constitue qu'une étape de l'expression génique.
Les protéines nouvellement synthétisées peuvent ensuite subir diverses modifications co- et post-traductionnelles qui participent à leur maturation, leur adressage et leur régulation.
- Les modifications co-traductionnelles surviennent pendant la synthèse de la chaîne polypeptidique et comprennent notamment la N-glycosylation dans le réticulum endoplasmique ou l'insertion membranaire de certaines protéines.
- Les modifications post-traductionnelles interviennent après la libération de la protéine et comprennent notamment des phosphorylations, des acétylations, des O-glycosylations ou des ubiquitinations, qui modulent la structure, la localisation, la stabilité et la fonction des protéines.
Les modifications post-traductionnelles sont étudiées dans un chapitre spécifique.
BiochimieChimie organiqueBioénergétiqueAcides nucléiquesNucléotidesADNARNStructure de l'ARNARN messager (ARNm)ARN de transfert (ARNt)ARN ribosomiques (ARNr)ARN régulateurs et fonctionnelsMaturation des ARNÉpissageÉditionÉpitranscriptomeTraductionSurveillance des ARNDégradation des ARNChromatineNucléosomesHistonesChromosomesProtidesGlucidesLipidesEnzymesCoenzymesVitaminesHormonesComposés inorganiquesTransport membranaireMoteurs moléculairesVoies de signalisation