


Cycle cellulaire
Cyclines : structure et classification
- Biologie cellulaire et moléculaire
- Constituants de la cellule
- Reproduction cellulaire
- Biochimie
- Transport membranaire
- Moteurs moléculaires

Les cyclines constituent une large famille de protéines régulatrices dont l’expression et la dégradation contrôlent des processus cellulaires essentiels, notamment le cycle cellulaire, la transcription, la différenciation et certaines voies de signalisation.
Les cyclines ont été nommées à l'origine parce que leur concentration varie de façon cyclique pendant le cycle cellulaire.
- Elles exercent leurs fonctions principalement en formant des complexes avec les kinases dépendantes des cyclines (CDK), modulant ainsi la phosphorylation de substrats spécifiques.
- Certaines cyclines, cependant, exercent des rôles indépendants des CDK, notamment dans la régulation de la dégradation protéique ou dans des voies de signalisation spécialisées.
Structure et activation
La structure primaire des cyclines est très différente les unes des autres, mais tous les membres de la famille sont caractérisés par une structure tridimensionnelle hautement conservée, dite cyclin fold.
Cette architecture repose sur deux domaines globulaires homologues, appelés boîtes à cycline (cyclin boxes), d’environ 100 résidus chacun, constitués majoritairement d'hélices α organisées selon un repliement antiparallèle (Insights on Structural Characteristics and Ligand Binding Mechanisms of CDK2 2015).
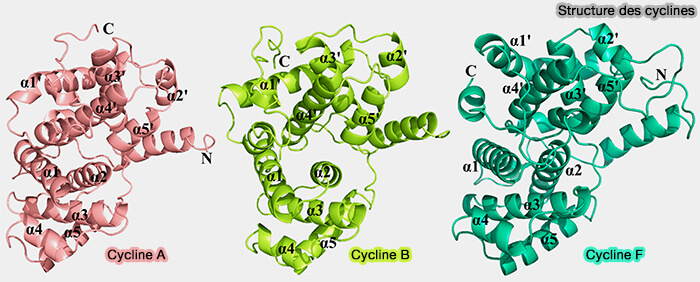
(Figure : vetopsy.fr d'après Li et coll)
Coeur structural : boîtes à cycline
1. Les deux boîtes à cycline sont reliées par une boucle inter-domaines flexible, qui confère à la protéine une certaine plasticité conformationnelle tout en conservant un cœur structural stable (The structure of cyclin E1/CDK2: implications for CDK2 activation and CDK2-independent roles 2005).
a. L’agencement tridimensionnel des deux domaines forme une surface interne concave hydrophobe composée principalement d’hélices α.
b. Cette surface constitue l’interface principale d’interaction avec la CDK, notamment par reconnaissance du motif PSTAIRE (ou de ses variantes proches) situé sur l’hélice αC de la kinase, et :
- permet l’association structurale entre cycline et CDK,
- explique l’activation conformationnelle de la kinase,
- fournit un cadre commun à toutes les cyclines.
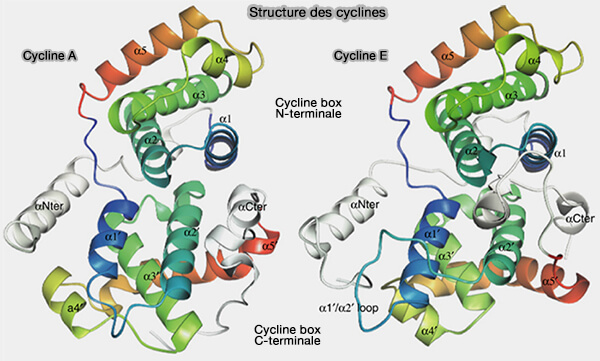
(Figure : vetopsy.fr d'après Honda et coll)
2. La première boîte à cycline, située à l’extrémité N-terminale du cœur structural, participe fortement à la formation de l’interface avec la CDK et contribue à la compatibilité générale entre partenaires cycline/CDK.
Elle contribue majoritairement à la formation de l’interface avec la CDK pour former un complexe actif et détermine en grande partie la spécificité de liaison à la CDK, donc quelle kinase sera activée.
3. La seconde boîte à cycline, localisée à l'extrémité C-terminale, complète cette interface et contribue à la stabilité globale du complexe.
Les différences fines d’interaction entre cyclines proviennent en grande partie de variations de résidus exposés à la surface, localisées dans des boucles et hélices périphériques du repliement, ainsi que dans les régions adjacentes au domaine C-terminal.
Ces variations modifient localement la topographie, la distribution des charges et l’accessibilité de certaines poches de surface, ce qui permet une diversification des interactions sans altérer l’architecture tridimensionnelle globale conservée.
Régions N- et C-terminales
En dehors du cœur constitué par les deux boîtes à cycline, les régions périphériques, en particulier les extrémités N-terminales et C-terminales, présentent une variabilité de séquence beaucoup plus importante selon les types de cyclines, ce qui contribue largement à leur spécificité fonctionnelle.
1. La région N-terminale et les segments périphériques associés peuvent contenir :
- des signaux d’import (NLS) ou d'export nucléaire (NES) régulant la localisation subcellulaire du complexe cycline/CDK comme CRM1-dépendant pour la cycline B,
- des motifs de destruction fréquemment portés par la région N-terminale, notamment des D-box, KEN-box, et/ou des régions PEST, impliqués dans la reconnaissance par les complexes de type APC/C (Anaphase-Promoting Complex/Cyclosome) ou SCF et dans la dégradation contrôlée au cours du cycle cellulaire.
2. La région C-terminale et les surfaces du cyclin fold associées comprennent :
- des motifs de recrutement des substrats, correspondant au site Cy (docking site) de la cycline, qui reconnaît le motif RXL (Cy-motif) présent dans de nombreux substrats des complexes cycline/CDK,
- les motifs ABBA (ou A-box docking motif), impliqués dans la reconnaissance préférentielle de certains substrats mitotiques par les cyclines A et B et contribuant à la hiérarchisation temporelle des phosphorylations.
3. De nombreux partenaires régulateurs reconnaissent les motifs portés par les régions N- et C-terminales ou par des surfaces périphériques du cyclin fold décrites ci-dessus, tels que: :
- des protéines adaptatrices, comme Chk1 et Chk2, les CKI (CDK inhibitors) p21CIP1 ou p27KIP1, ou encore Skp2, qui interagissent avec les complexes cycline/CDK, qui modulent l’état d’activation et la dynamique fonctionnelle des complexes,
- des cofacteurs régulateurs, comme Cdc25, Wee1 et Myt1 (PKMYT1), le complexe CAK (cycline H/CDK7/Mat1) ou Chk1/2, qui modulent l’état d’activation et la dynamique fonctionnelle des complexes,
- des composants de complexes de contrôle, notamment APC/C (via Cdc20 ou Cdh1), SCF (Skp1-Cullin-F-box), ainsi que des acteurs du transport nucléocytoplasmique comme CRM1 (exportine-1) ou les importines nucléaires.
Classification
Le classement des cyclines ne suit pas l’ordre alphabétique mais une logique fonctionnelle et chronologique.
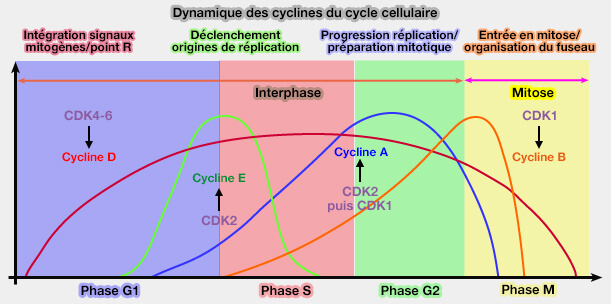
(Figure : vetopsy.fr)
1. Les cyclines du cycle cellulaire sont organisées selon leur séquence d’apparition et d’action (G1 → S → G2 → M),
2. Les cyclines transcriptionnelles sont disposées selon leur hiérarchie fonctionnelle, allant des régulateurs globaux (activation des CDK, initiation transcriptionnelle) aux fonctions les plus spécialisées (épissage).
3. Les cyclines différenciatives sont présentées selon leur degré de spécialisation physiologique et classées selon leur importance physiologique et leur degré de spécialisation, de la réponse au stress aux rôles développementaux les plus spécifiques.
| Cycline | CDK associée(s) |
Phase ou fonction principale |
Rôle biologique majeur |
Particularités structurales/ localisation |
|---|---|---|---|---|
| Cyclines liées au cycle cellulaire (classement fonctionnel et chronologique) |
||||
| Cycline D (D1, D2, D3) |
CDK4/ CDK6 |
G1 |
|
|
| Cycline E |
CDK2 | Fin G1/ Transition G1-S |
|
|
| Cycline A |
CDK2/ CDK1 |
S/G2 |
|
|
| Cycline F |
Indépendante des CDK |
|
|
|
| Cycline B (B1, B2) |
CDK1 | G2/M ➞ mitose |
|
|
| Cyclines transcriptionnelles et co-transcriptionnelles (classement par hiérarchie fonctionnelle) |
||||
| Cycline H |
CDK7 | Tout le cycle |
|
|
| Cycline C |
CDK8/ CDK19 |
Transcription
|
|
|
| Cycline K |
CDK12/ CDK13 |
Transcription et stabilité de l'ARN |
|
|
| Cycline T (T1, T2) |
CDK9 | Transcription (P-TEFb) |
Phosphoryle le CTD de l'ARN polymérase II pour activer l'élongation transcriptionnelle |
|
| Cycline L |
CDK11 (p110/p58) |
|
|
|
| Cyclines différenciatives ou tissulaires (classement par importance fonctionnelle et ancienneté évolutive) |
||||
| Cycline G1/G2 |
CDK2/ CDK5 |
Réponse au stress |
|
|
| Cycline I (Ccni) |
CDK5 | Régulation post-mitotique neuronale |
|
|
| Cycline I2 (Ccni2) |
CDK5 | Différenciation neuronale |
|
|
| Cycline O |
CDK2 ? |
|
|
|
| Cycline Y (CCNY) |
CDK14- CDK18 |
Signalisation Wnt |
|
|
| Cycline J (Ccnoj) |
Fonction en cours d'étude |
Développement/ différenciation ovarienne |
|
|
| Cycline P (CNPP) |
Fonction en cours d'étude |
Différenciation/ Régulation de l’entrée en cycle |
|
|
Rôles physiologiques des cyclines liées au cycle cellulaire
Biologie cellulaire et moléculaireConstituants de la celluleReproduction cellulaireMatériel génétiqueFuseauCycle cellulaireInterphasePhase G1Phase SPhase G2MitoseProphasePrométaphaseMétaphaseAnaphaseTélophaseCytokinèseAbcissionMéioseRégulation du cycle cellulaireCyclinesCDKComplexes Cyclines/CDKPoints de contrôles (checkponts)Enzymes mitotiquesMoteurs mitotiquesComplexes SMC (condensines et cohésines)BiochimieTransport membranaire Moteurs moléculairesVoies de signalisation