


Lipides
Lipides complexes
: phospholipides
Glycérophospholipides
Cardiolipine (diphosphatidylglycérol) : vue d'ensemble
- Biochimie
- Chimie organique
- Bioénergétique
- Composition de la matière vivante
- Composés organiques
- Protides
- Acides nucléiques
- Glucides
- Lipides
- Vue d'ensemble des lipides
- Acides gras
- Classification des lipides
- Lipides vrais
- Composés à caractère lipidique, lipoïdes ou lipides insaponifiables
- Phospholipides
- Cholestérol
- Glycérides
- Trafic non vésiculaire des lipides et LTP
- Digestion et absorption entérocytaire des lipides
- Lipoprotéines
- Gouttelettes lipidiques (LD)
- Coenzymes
- Hormones
- Composés inorganiques
- Composés organiques
- Transport membranaire
- Moteurs moléculaires
- Voies de signalisation

La cardiolipine (diphosphatidylglycérol), chez les eucaryotes est un composant phospholipidique essentiel des membranes mitochondriales.
Le terme de " cardioline " provient du fait qu'elle a été découverte dans le cœur des animaux, où elle est particulièrement abondante, mais elle peut être trouvée dans les mitochondries de tous les tissus, mais aussi chez les procaryotes.
| Glycérophospholipides (forment les phospholipides avec les sphingophospholipides) |
|
|---|---|
| Azotés | Non azotés |
| Lysoglycérophospholipide | |

La cardiolipine (CL) est décrite de manière détaillée dans : Cardiolipin (Diphosphatidylglycerol) de LipidWeb.
Vue d'ensemble de la cardiolipine
1. La cardiolipine (CL) est retrouvée dans les organites dont la fonction est de générer un potentiel électrochimique pour transporter un substrat et synthétiser l'ATP.

La cardiolipine est localisée presque exclusivement dans la membrane mitochondriale interne (IMM) des mitochondries des eucaryotes, i.e. elle est indispensable au fonctionnement optimal de nombreuses enzymes impliquées dans le métabolisme énergétique mitochondrial.
2. La cardiolipine est présente en quantité importante :
- dans le coeur, i.e. environ 10% des phospholipides et 20% des phospholipides de ses membranes mitochondriales, 25% sur l'IMM, 4% sur l'OMM.
- dans les muscles squelettiques, i.e. à forte demande d'énergie.
Les autres organes en contiennent des quantités plus faibles.
3. La cardiolipine est considérée comme un " lipide de signature " des mitochondries, i.e. sa présence dans les membranes permet de reconnaître que c'est une mitochondrie.
Elle est aussi présente dans les membranes peroxysomales.
Structure de la cardiolipine
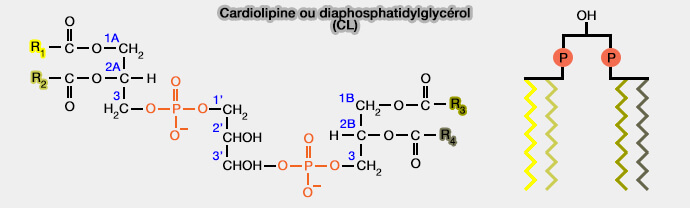
1. La cardiolipine (1,3‑bis(sn‑3'-phosphatidyl)-sn-glycérol), seul phospholipide que l'on peut considérer à structure dimérique.
a. Les quatre groupes acyle ($\ce{CH2-O-C(=O)-R}$) rend la molécule très complexe.
- Cependant, dans la plupart des tissus animaux, la cardiolipine contient des chaînes acyles grasses à 18 carbones avec 2 liaisons insaturées sur chacune d'elles, comme l'acide linoléique (18:2 n-6),
- Cette configuration de la chaîne acyle est une exigence structurelle importante pour la haute affinité de CL pour les protéines de la membrane mitochondriale interne (IMM).
(Figure : vetopsy.fr)
b. Les deux groupes phosphates, i.e. appelé 1' et 3' par rapport au glycérol central, possède chacun une charge négative.
- Contrairement à ce que l'on pensait précédemment, les pH d'ionisation sont proches très différent, i.e. pK1, 2,15 et pK2, 3,15, au lieu de 7,5.
L'acidité plus faible du second phosphate serait due à la formation d'une liaison hydrogène intramoléculaire stable avec le groupe 2'-hydroxyle du glycérol central.
- Dans des conditions physiologiques normales, à pH d'environ 7, la cardiolipine porte une double charge négative, ce qui est très utile pour la phosphorylation oxydative.
2. Comme le groupe de tête forme une structure compacte, la zone du groupe de tête est assez petite, mais compacte par rapport à la grande région de queue flexible composée de 4 chaînes acyle.
En raison de cette structure unique, la cardiolipine peut former des états micellaires, lamellaires et hexagonaux dans des dispersions aqueuses, en fonction du pH et de la force ionique ( cardiolipine et membranes)
cardiolipine et membranes)
Synthèse de la cardiolipine
Vous pouvez lire : Biosynthesis, remodeling and turnover of mitochondrial cardiolipin (2017).
Synthèse générale
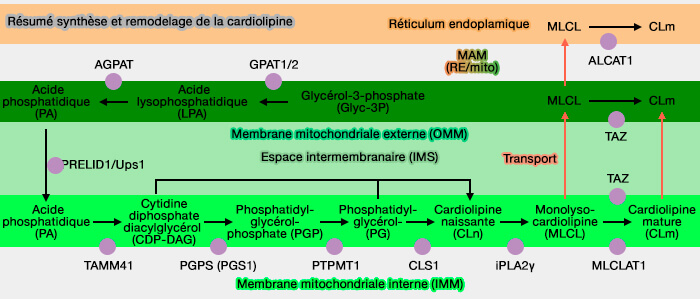
La voie de synthèse de la cardiolipine est commune aux glycérophospholipides et passe par l'acide phosphatidique (PA) qui est (voie du glycérol-3-phosphate (glyc-3P) :
- soit importé du réticulum endoplasmique (RE) et transporté vers la membrane mitochondriale externe (OMM) par des complexes protéiques spécifiques comme le complexe ERMES (ER-Mitochondria Encounter Structure).
- soit synthétisé directement sur la face externe de la membrane mitochondriale externe (OMM).
Dans les mitochondries, PA est transféré dans sur la face interne de la membrane mitochondriale interne (IMM) par Ups1/PRELID1.
- La suppression d'Ups1 diminue les niveaux de CL, i.e. la restauration est effectuée par la suppression simultanée de Ups2 (Ups1p and Ups2p antagonistically regulate cardiolipin metabolism in mitochondria 2009).
- Mdm35/TRIAP1 facilite l'import mitochondrial des Ups1 et les protège contre la protéolyse.

(Figure : vetopsy.fr d'après Miliara et coll)
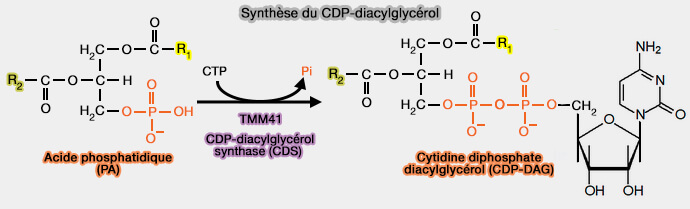
1. L'acide phosphatidique (PA) est transformé en cytidine diphosphate diacylglycérol (CDP-DAG).
$\ce{CTP + acide phosphatidique}$ $\leftrightharpoons$ $\ce{CDP-DAG + diphosphate}$
a. Dans les mitochondries animales, cette réaction est catalysée par TAMM41 (Translocator Assembly and Maintenance Mitochondrial 41), TAM41 chez les levures, localisée à l'IMM face à la matrice (Tam41 Is a CDP-Diacylglycerol Synthase Required for Cardiolipin Biosynthesis in Mitochondria 2013 et Biallelic variants in TAMM41 are associated with low muscle cardiolipin levels, leading to neonatal mitochondrial disease 2022).
(Figure : vetopsy.fr)
b. Sur la surface cytoplasmique du réticulum endoplasmique (RE), cette réaction est catalysée par la phosphatidate cytidylyltransférase ou CDP-diacylglycérol synthase (CDS), i.e. EC 2.7.7.41.
Les CDS n'a aucune relation structurelle avec TAMM41 et comprend deux isoformes, CDS2 ubiquitaire et CDS1 dans le coeur.
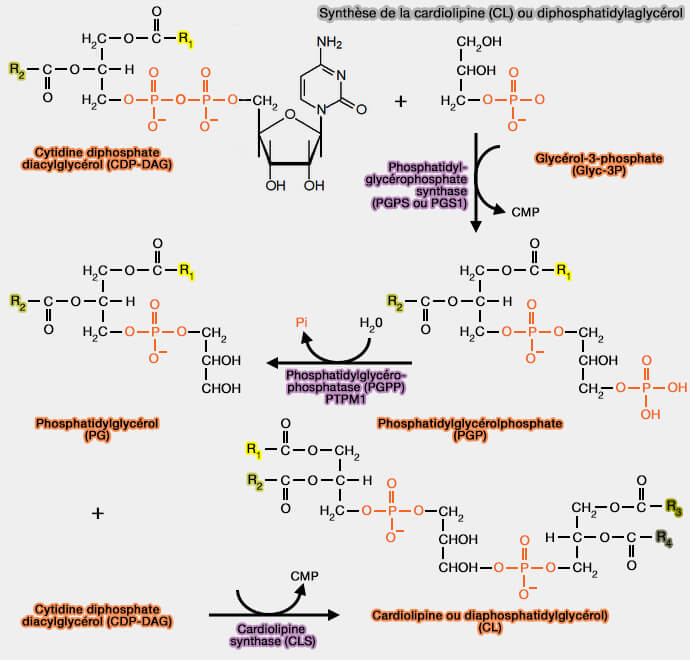
2. CDP-DAG interagit avec le glycérol-3-phosphate (Glyc-3P) pour former le phosphatidylglycérolphosphate (PGP ou 3-sn-phosphatidyl-1'-sn-glycerol 3'-phosphoric acide), réaction catalysée par la phosphatidylglycérophosphate synthase (PGPS ou PGS1) ou CDP-diacylglycérol—glycérol-3-phosphate 3-phosphatidyltransférase (EC 2.7.8.5).
(Figure : vetopsy.fr)
3. Le phosphatidylglycérolphosphate (PGP) est transformé en phosphatidylglycérol (PG) par la PTPMT1 ou PTP [(Protein Tyrosine Phosphatase) localized to the MiTochondrion 1], une phosphatidylglycérophosphatase PGPP), i.e. EC 3.1.3.27 (Structural and functional analysis of PTPMT1, a phosphatase required for cardiolipin synthesis 2011).
4. Le phosphatidylglycérol (PG) réagit avec la CDP-DAG pour former la cardiolipine (diphosphatidylglycérol), réaction catalysée par la cardiolipine (ou diphosphatidylglycérol) synthase (CLS), une phosphatidyl transférase (EC 2.7.8.41) .

La cardiolipine nouvellement synthétisée ou CLn, i.e. cardiolipine naissante, est une CL immature, caractérisée par des chaînes acyles saturées asymétriques et de longueur variable autour du groupe de tête glycérol.
(Figure : vetopsy.fr)
Remodelage de la cardiolipine
La cardiolipine nouvellement synthétisée (CLn) subit un remodelage, un processus de désacylation et de réacylation, pour devenir une cardiolipine mature.
Enzymes mises en jeu
Dans les mitochondries, la cardiolipine est remodelée, mais le processus est plus complexe que pour les autres phospholipides au vu de ces quatre chaînes acyle.
1. La majeure partie de ce remodelage fait appel à la tafazzine, ancrée à la surface externe de la membrane mitochondriale interne (IMM) par un segment hydrophobe (The Enzymatic Function of Tafazzin 2006 et Mechanism for Remodeling of the Acyl Chain Composition of Cardiolipin Catalyzed by Saccharomyces cerevisiae Tafazzin 2016).
2. Les mammifères possèdent deux autres enzymes capables de réacyler la MLCL, (Cardiolipin remodeling: a regulatory hub for modulating cardiolipin metabolism and function 2016) :
- ALCAT1 (acyl-CoA : lysocardiolipine acyltransférase-1), associée à la membrane du réticulum endoplasmique (RE) associée aux mitochondries (MAM),
- MLCLAT1 (MLCL AcylTransférase 1), à la surface interne de la membrane mitochondriale interne (IMM).
Remarque : les localisations exactes des enzymes peuvent varier suivant les auteurs.
Modes d'action
1. Quelle que soit l'enzyme, le remodelage de la cardiolipine commence par une désacylation ( cycle de Lands).
- Tout d'abord, un acide gras en sn-2 est libéré par la phospholipase A2 (PLA2), comme pour les autres phospholipides pour former, ici, une monolysocardiolipine (MLCL).
- Cette PLA2 est la PNPLA8, également appelée phospholipase A2 γ indépendante du calcium (iPLA2γ), qui élimine une chaîne acyle grasse saturée du CL.
2. Cette désacylation est suivie par une réacylation par une acyltransférase pour introduire une chaîne d'acyle gras insaturé.
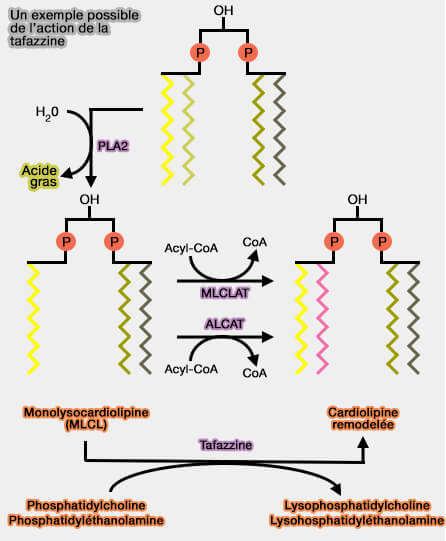
(Figure : vetopsy.fr d'après Saric et coll)
a. La taffazine (TAZ), i.e. acyltransférase coenzyme-A-indépendante, localisée sur la surface externe de la membrane mitochondriale interne (IMM).
- TAZ est une transacylase, effectuant un échange de chaîne acyle entre la cardiolipine et les phospholipides (PL), principalement la phosphatidylcholine (PC), ou, plutôt la plasménylcholine pour remplacer séquentiellement les chaînes acyles aux quatre positions acyles.
- La carence en tafazzine entraîne une diminution de la teneur en cardiolipine et une augmentation de celle des MLCL.
b. ALCAT1 (MAM) et MLCLAT1 (IMM) catalysent le remodelage de la cardiolipine en ajoutant de longues chaînes d'acyles gras polyinsaturés.
ALCAT1 et MLCLAT1 utilisent la chaîne acyle de l'acyl-CoA pour assurer la réacylation de MLCL.
3. Le remodelage déplace la composition des acyles vers l'insaturation, bien la spécificité de substrat soit relativement faible, surtout pour la tafazzine.
L'environnement membranaire pourrait être une source de spécificité dans les réactions.
4. Les quatre chaînes d'acyle gras de la cardiolipine dans les tissus métaboliques, y compris le cœur, sont dominées par l'acide linoléique (18:2 n-6), appelé tétralinoléoyl CL (TLCL), ce qui indique que les enzymes impliquées dans le remodelage dans ces tissus peuvent utiliser l'acide linoléique comme substrat majeur (Cardiolipin remodeling by ALCAT1 links hypoxia to coronary artery disease by promoting mitochondrial dysfunction 2021).
- Par exemple, elle génère la tétralinoléoyl cardiolipine (L4-CL), le principal constituant de la cardiolipine dans le cœur et sa forme s'adapte particulièrement bien à la géométrie des monocouches à courbure négative, de sorte que cette structure pourrait être favorisée.
5. La tafazzine est impliquée dans le syndrome de Barth ( cardiolipine et pathologies).
Transport de la cardiolipine dans les mitochondries
1. Le transport de la cardiolipine dans l'IMM et l'OMM serait assurée par trois enzymes différentes (Mitochondrial cardiolipin/phospholipid trafficking: The role of membrane contact site complexes and lipid transfer proteins 2014) :
- phospholipase scramblase,
- mitochondrial creatine kinase ou MtCK (Mitochondrial Creatine Kinase Binding to Phospholipid Monolayers Induces Cardiolipin Segregation 2009),
- nucleoside polyphosphate kinase ou NDPK-D/Nm23-H4 (NDPK-D (NM23-H4)-mediated externalization of cardiolipin enables elimination of depolarized mitochondria by mitophagy 2016).
2. Les activités de ces enzymes sont également régulées par la cardiolipine et nécessitent une agrégation de protéines qui favorise la formation de groupement et de domaines membranaires enrichies en cardiolipine.
Ces domaines enrichis en CL jouent un rôle important dans la régulation de la structure et de la morphologie de la membrane mitochondriale, ainsi que dans l'agrégation des protéines (Cardiolipin Clusters and Membrane Domain Formation Induced by Mitochondrial Proteins 2007).
Fonctions de la cardiolipine
BiochimieChimie organiqueBioénergétiqueProtidesGlucidesLipidesAcides grasLipides vraisLipides vrais simplesLipides vrais complexesLipoïdesCholestérolGlycéridesPhospholipidesGlycérophospholipidesSphingophospholipidesTrafic non vésiculaire et LTPDigestion et absorption entérocytaire des lipidesLipoprotéinesApolipoprotéinesGouttelettes lipidiques (LD)EnzymesCoenzymesVitaminesHormonesComposés inorganiquesTransport membranaireMoteurs moléculairesVoies de signalisation