Les cyclines F et G régulent indirectement le cycle cellulaire en contrôlant la stabilité des protéines, les checkpoints et la réponse au stress plutôt que l’activation des CDK.
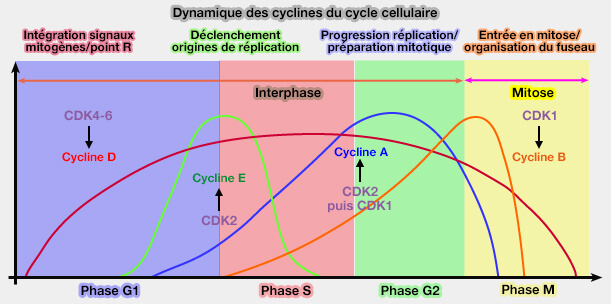
Dynamique des cyclines du cycle cellulaire
(Figure : vetopsy.fr)
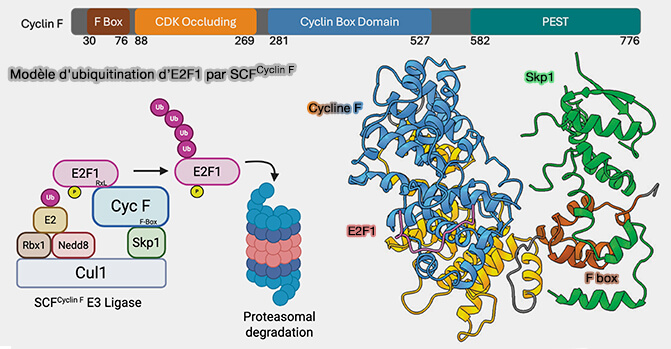
Cycline F
La cycline F (CCNF) est une cycline atypique qui ne s’associe à aucune CDK (Cyclin-Dependent Kinases), malgré la présence de deux boîtes à cycline caractéristiques de sa famille.
1. La cycline F fonctionne comme protéine F-box, cas particulier de la famille FBXO, formant le complexe SCFcycline F, auquel elle confère des rôles essentiels dans :
la transition G1 ➞ S, en assurant l’extinction coordonnée de Cdh1,
le contrôle du licensing et la prévention du re-licensing des origines pendant la phase S,
la protection de l’intégrité génomique par la dégradation de facteurs impliqués dans la réplication, les centrosomes et la dynamique du fuseau.
APC/C et SCFcycline F
(Figure : vetopsy.fr d'après Michael et Taylor)
APC/CCdh1 dégrade la cycline F de l’anaphase B jusqu’à la phase G1 précoce, pour empêcher toute accumulation prématurée de régulateurs pro-S.
SCFcycline F dégrade Cdh1 en fin de G1 et en phase S, ce qui permet l’accumulation des protéines nécessaires à l’entrée et à la progression en phase S.
Cette alternance stricte stabilise le G1, autorise la montée des signaux pro-S, et synchronise l’articulation APC/C ➞ SCF au cours du cycle.
3. SCFcycline F cible plusieurs substrats clés impliqués dans la régulation du protéome nucléaire et la stabilité de nombreuses protéines, régulateurs du cycle et facteurs d’intégrité génomique comme :
RRM2 (Ribonucléotide Réductase M2), sous-unité essentielle de la ribonucléotide réductase responsable de la production des dNTP (désoxyribonucleoside triphosphates), substrats directs des ADN polymérases, qui est ciblée pour dégradation par SCFcycline F en fin de S afin de réduire le pool de nucléotides, prévenir le stress réplicatif et limiter l’instabilité génomique.
NUSAP1 (Nucleolar and spindle associated protein 1), facteur de dynamique des microtubules mitotiques dont
la quantité est régulée par la cycline F pour protéger la stabilité du fuseau,
SLBP (Stem-Loop Binding Protein), facteur de maturation des ARNm d’histones, dont la dégradation coordonne la disparition du programme histone en fin de phase S,
CDC6 (Cell Division Cycle 6), composant essentiel du complexe de pré‑réplicatif (pré-RC), dont l’ubiquitination limite le re-licensing en phase S et garantit l’unicité de la réplication.
Ubiquitination d'E2F par SCFcycline F
(Figure : vetopsy.fr d'après Ngoi et coll)
Cycline G
Spécificités fonctionnelles de la cycline G
1. La cycline G1/G2 est une cycline atypique, distincte des cyclines canoniques du cycle cellulaire.
Son association avec CDK2 ou CDK5 peut être observée dans certains contextes, mais elle est faible, indirecte et contextuelle, et ne constitue pas un mécanisme de contrôle des transitions G1/S ou G2/M.
La cycline G n’active pas de complexes cycline/CDK capables de piloter la progression du cycle, et ne fonctionne pas comme un effecteur de checkpoint.
Son rôle doit être compris comme celui d’un modulateur fin de certaines réponses cellulaires, sans capacité propre à imposer un ralentissement du cycle comparable aux mécanismes classiques (p21, Chk1, ATM/ATR).
2. Ces fonctions contextuelles incluent notamment :
une modulation de l’activité de CDK5 dans certains contextes neuronaux ou en réponse au stress,
une interaction ponctuelle avec CDK2, susceptible d’influencer localement certains complexes nucléaires sans impact global sur la dynamique du cycle,
une participation à la régulation de l’axe p53-PP2A-MDM2, qui ajuste l’intensité et la durée de la réponse au stress sans bloquer directement la progression du cycle.
Rôle dans le stress
Sa fonction principale est la modulation de la réponse au stress cellulaire via les voies p53-dépendantes, contribuant à l’équilibre entre réparation de l’ADN, arrêt transitoire du cycle et survie.
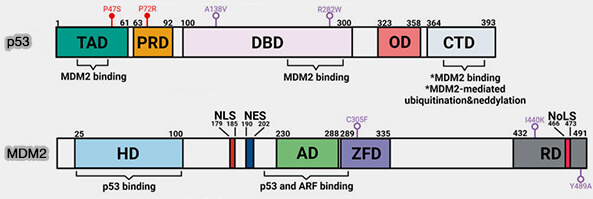
Structure de p53 et MDM2
(Figure : vetopsy.fr adaptée d'après Kung et Weber)
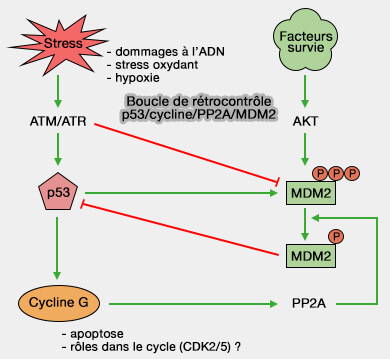
1. Les stress (dommages à l’ADN, stress oxydant, hypoxie) activent ATM/ATR et :
favorisent l’activation de p53,
Boucle de rétrocontrôle de p53
(Figure : vetopsy.fr)
limitent l’activité de MDM2, une ligase E3 de la famille RING et inhibiteur majeur de p53.
Remarque : à l’inverse, les signaux de survie via Akt activent MDM2 par phosphorylation, renforçant l’inhibition de p53.
2. L’expression de la cycline G est activée par p53.
La cycline G ne constitue pas un effecteur direct des checkpoints du cycle, mais agit comme modulateur de la dynamique de la réponse p53.
3. La cycline G, via PP2A, ajuste cet équilibre et participe à la régulation fine entre arrêt du cycle, réparation et apoptose.
Elle favorise la déphosphorylation de MDM2, modulant son activité et instaurant une boucle de rétrocontrôle :
p53 ➞ cycline G ➞ PP2A ➞ MDM2 ➞ p53.
Autres membres de la famille des cyclines
Plusieurs autres cyclines existent chez les eucaryotes, notamment les cyclines C, K, T, L, Y et certaines cyclines orphelines, principalement impliquées dans :