Biologie cellulaire
Mitochondries : génome
Structure et organisation de l'ADN mitochondrial
Sommaire
En construction
Le génome mitochondrial est un ADN circulaire d'origine bactérienne, transmis principalement par la lignée maternelle et codant un nombre limité de gènes indispensables à la fonction des mitochondries.
Les mitochondries possèdent leur propre matériel génétique, distinct du génome nucléaire, dont l'organisation compacte et le mode d'expression particulier reflètent leur origine endosymbiotique.
Bien que ne représentant qu'une faible fraction du patrimoine génétique cellulaire, ce génome est indispensable à la production d'énergie et à de nombreuses fonctions mitochondriales.
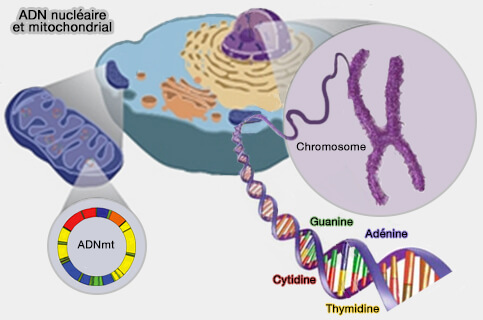
ADN mitochondrial et nucléaire
(Figure : vetopsy.fr)
ADN mitochondrial humain
L'ADN mitochondrial humain (ADNmt) est une molécule circulaire bicaténaire de 16 569 paires de bases présente en de multiples copies dans chaque cellule.
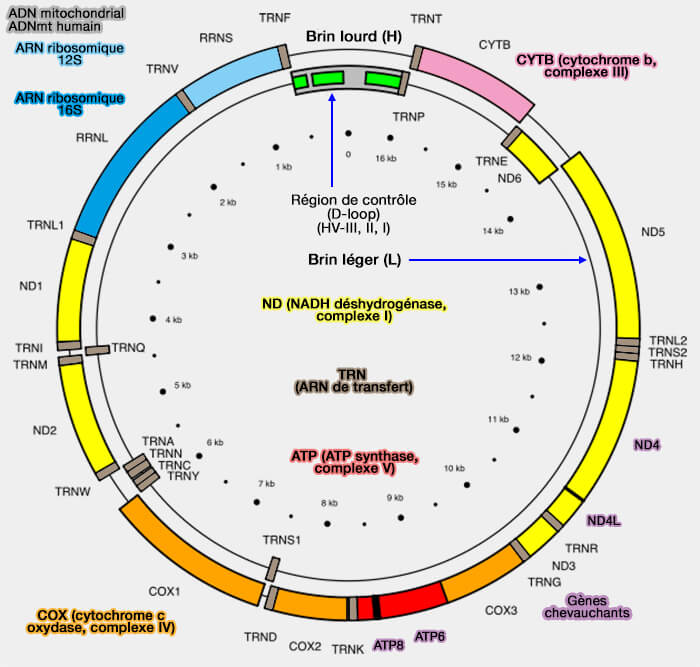
Gènes mitochondriaux
Héritage d'un ancien génome bactérien, l'ADNmt est beaucoup plus compact que le génome nucléaire, contient très peu de séquences intergéniques et est constitué :
d'un brin lourd (H pour Heavy), riche en guanines, qui code 28 gènes,
d'un brin léger (L pour Light), riche en cytosines qui en code 9.
Carte de l'ADN mitochondrial humain
(Figure : vetopsy.fr adaptée d'après Emmanuel Douzery)
Remarque : plusieurs études ont proposé l'existence de peptides supplémentaires codés par des cadres de lecture ouverts (ORF) présents dans l'ADNmt.
Le plus étudié est l'humanine, un peptide dérivé de la région du gène mitochondrial MT-RNR2 (ARNr 16S), auquel ont été attribuées des fonctions cytoprotectrices.
3. Comparé au génome nucléaire, le génome mitochondrial possède plusieurs particularités.
Il est présent en de nombreuses copies par cellule.
Il est transmis principalement par la lignée maternelle et présente un taux de mutation généralement plus élevé.
Son organisation compacte, la quasi-absence d'introns et son expression génique particulière témoignent de son origine bactérienne et de son évolution spécifique.
Région de contrôle de l'ADNmt
(Figure : vetopsy.fr d'après Falkenberg)
les séquences TAS (Termination-Associated Sequences), associées à la régulation de la région D-loop,
trois blocs de séquences conservées, CSB I, CSB II et CSB III (Conserved Sequence Blocks), impliqués dans la régulation de la transcription et de la réplication.
1. Les blocs CSB correspondent à de courtes séquences d'ADN hautement conservées au cours de l'évolution qui, contrairement aux promoteurs ou aux origines de réplication, ne constituent pas des sites d'initiation, mais des éléments régulateurs qui modulent le déroulement de la transcription et de la réplication dans la région de contrôle.
Les fonctions précises de CSB I et CSB III restent en revanche moins bien établies et participeraient à l'organisation de la région de contrôle et au bon déroulement des mécanismes de transcription et de réplication, sans que leur rôle moléculaire exact soit encore complètement élucidé.
2. La région de contrôle contient fréquemment un court troisième brin d'ADN, appelé ADN 7S, synthétisé à partir de l'origine de réplication OH.
Lorsque ce brin reste associé à la molécule d'ADN mitochondrial après sa synthèse, il déplace localement le brin lourd et forme une structure à trois brins appelée D-loop (Displacement loop).
Toutes les molécules d'ADN mitochondrial ne possèdent cependant pas un ADN 7S, dont le rôle exact reste encore discuté.
3. a région de contrôle comprend également les régions hypervariables HV-I, HV-II et HV-III, qui accumulent de nombreuses variations de séquence entre les individus et constituent des marqueurs largement utilisés en phylogénie, en anthropologie et en médecine légale.
Origine du génome mitochondrial
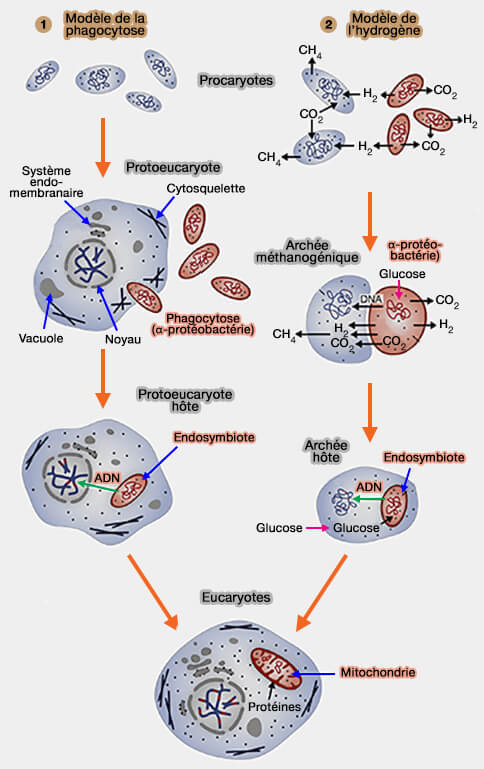
Le génome mitochondrial est considéré comme le vestige d'un ancien génome bactérien acquis lors d'un événement d'endosymbiose survenu au cours de l'évolution des eucaryotes.
b. Cette endosymbiose aurait eu lieu il y a environ 1,5 à 2 milliards d'années, dans un contexte d'enrichissement progressif de l'atmosphère en oxygène
2. L'endosymbiose, du grec endon "dans ", syn " ensemble " et biosis " vivant " implique la coopération mutuellement bénéfique entre deux organismes vivants, où l'un est contenu dans l'autre.
a. Ce génome, au cours de cette endosymbiose, s'est réduit drastiquement, car l'endosymbiote a transféré la plus grande partie de ses gènes à l'hôte (transfert des gènes et hypothèse CoRR).
b. Malgré cette réduction considérable, le génome mitochondrial a conservé plusieurs caractéristiques héritées de son origine bactérienne, notamment sa structure généralement circulaire, son organisation compacte, sa transcription polycistronique et sa propre machinerie de traduction.
Remarque : l'ADN mitochondrial est circulaire chez l'homme et la plupart des métazoaires, mais certaines algues, certains ciliés ou certains cnidaires possèdent des génomes mitochondriaux linéaires ou plus complexes.
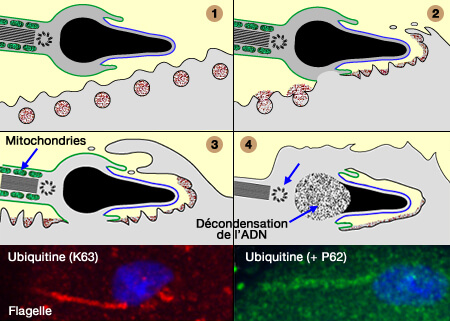
Elle agit en coopération avec l'autophagie maternelle et les mécanismes dépendants de l'ubiquitine pour favoriser l'élimination des mitochondries paternelles par les protéasomes.
La perte de CPS-6 retarde la dégradation des membranes internes mitochondriales, perturbe l'autophagie et augmente la létalité embryonnaire, soulignant l'importance de ce mécanisme pour le développement normal.
2. La transmission essentiellement maternelle de l'ADNmt a permis de définir le concept d'Ève primordiale ou Ève mitochondriale, ancêtre féminine hypothétique dont dérivent tous les ADN mitochondriaux humains actuels.
Remarque : le concept d'Ève mitochondriale est parfois rapproché de celui d'Adam chromosomique Y, qui correspond à l'ancêtre paternel commun le plus récent des chromosomes Y humains actuels, bien que ces deux concepts soient totalement indépendants ( goulot d'étranglement de population).
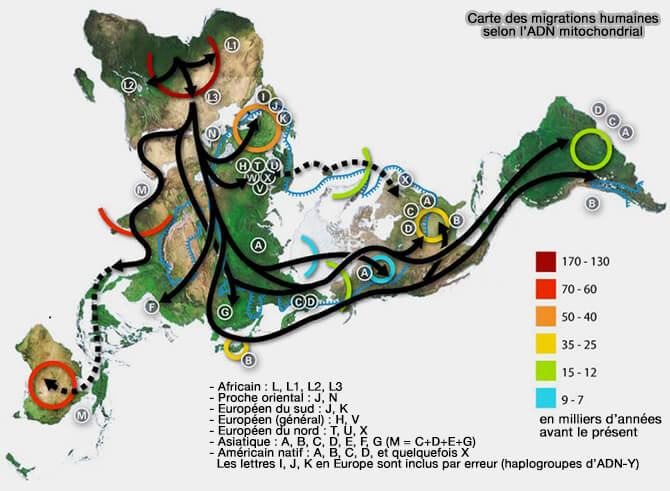
Carte des migrations humaines selon l’ADN mitochondrial
(Figure : vetopsy.fr d'après Avsa)
3. Les mitochondries se multiplient par fission et forment, avec les mécanismes de fusion mitochondriale, un réseau dynamique appelé chondriome, capable de s'adapter aux besoins métaboliques de la cellule.
Cette dynamique mitochondriale participe également au contrôle de la qualité des mitochondries et à l'élimination des génomes mitochondriaux porteurs de mutations délétères ( contrôle de la qualité mitochondriale et fusion).
Chez les mammifères, les cas sont exceptionnels comme chez la souris, les clones de moutons ou de bovins.
Ainsi, bien que la transmission maternelle constitue la règle générale chez les animaux, des exceptions rares ou des systèmes particuliers d'hérédité mitochondriale peuvent être rencontrés.




{kind=link}
 assemblage des complexes respiratoires et coopération noyau-mitochondrie),
assemblage des complexes respiratoires et coopération noyau-mitochondrie),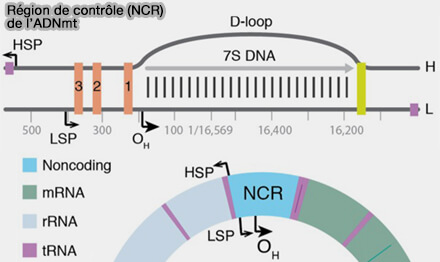
{kind=link}

