Les protéine kinases sont des enzymes spécifiques du groupe des transférases catalysant les réactions de phosphorylation par l'ajout d'un groupe phosphate à des protéines à partir de l'ATP.
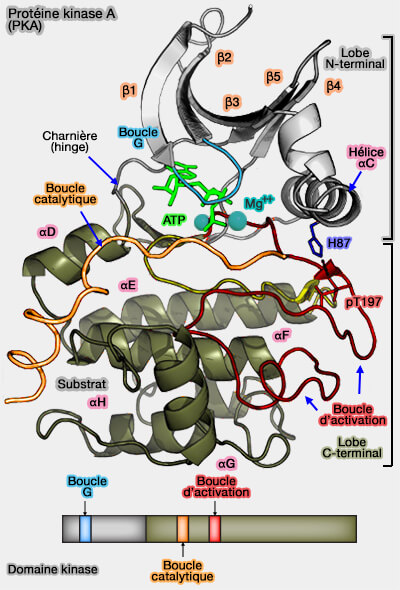
d'une fente catalytique située entre les deux lobes.
Remarque : les deux lobes sont souvent considéré comme des plis Rossmann-like.
Lobe N-terminal
Le petit lobe N-terminal, résidus 40-119, est formé par un feuillet β (β sheet) antiparallèle à 5 brins (β1 à β5) et une petit domaine hélicoïdal α à 3 hélices (αA-αC).
L'extrémité N-terminale (Gly1) peut être myristolisée pour augmenter son affinité membranaire.
Boucle G
La boucle G, riche en glycine (G50, G52 et G55) relie β1 et β2 et forme une sorte de couvercle au-dessus de l'entrée dans la cavité catalytique.
1. Dans la conformation fermée, la boucle G recouvre l'ATP le protégeant ainsi du solvant, et forme la première partie de l'ancrage de l'ATP.
G52 régule la position de Ser53 qui forme une liaison hydrogène avec le γ phosphate,
Phe54 et Gly55 ancrent le β phosphate.
2. La boucle G se termine avec Val57 (β2), qui avec Ala70 forme la partie supérieure de l'interstice hydrophobe contenant l'hétérocycle d'adénine de l'ATP.
Brin β3
1. Une partie du brin β3 forme la deuxième partie de l'ancrage de l'ATP.
Lys72, dans la clef catalytique, interagit directement avec les α et β phosphate de l'ATP.
2. D'autres éléments sur le lobe C coordonnent aussi l'ATP.
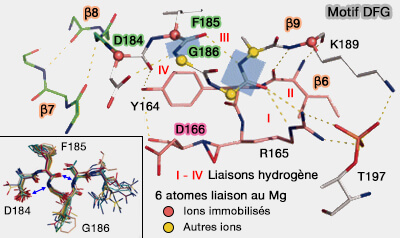
Lys168 de la boucle catalytique se lie au γ phosphate de l'ATP et sa position est déterminée par les interactions de Thr197–Arg165–Asp166–Thr201–Lys168.
Motif DFG
(Figure : vetopsy.fr d'après Kornev et coll)
Mg1 coordonne les α- et γ-phosphates de l'ATP par la liaison avec Asn171.
Mg2 coordonne les β- et γ-phosphates d'ATP. par la liaison avec dAsp184.
Hélice αC
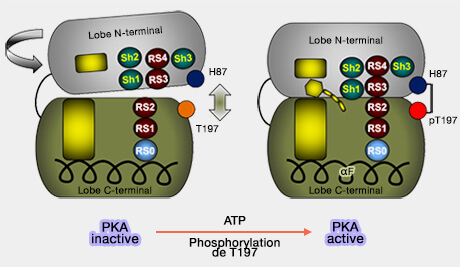
L'hélice αC, placée entre les deux lobes de la PKA près du segment d'activation, est mobile et dépend de la phosphorylation, de la liaison de l'ATP et de l'assemblage des spines.
La conformation de la PKA est fermée par une liaison entre His87 et pThr197 de la boucle d'activation.
Région charnière
Le lien flexible, résidus 120-127, est situé entre les deux lobes au fond de la fente catalytique avec l'hélice αD.
1. Glu127 ancre le groupe 1′-OH du ribose, le groupe 2'-OH étant fixé par Glu170 de la boucle catalytique.
2. Glu121 et Val123 reconnaissant les atomes d'azote de noyau purique de l'adénine.
Les spines C et R de PKA
(Figure : vetopsy.fr d'après Meharena)
Le groupe carbonyle (C=O) de Glu121 forme une liaison hydrogène avec le groupe NH2 de l'adénine.
Le groupe amide de Val123 connecte la purine au lien.
L'adénine est également coordonnée par une liaison hydrogène avec Thr183.
3. L'ATP, se liant simultanément avec les deux lobes, les maintient ensemble et fait tourner l'enzyme dans une conformation fermée.
En l'absence d'ATP, la boucle d'activation et les spines sont déformées, déstabilisant la molécule.
Les lobes N et C peuvent se déplacer l'un par rapport à l'autre en raison de la liaison flexible (conformation ouverte).
La boucle de liaison au magnésium sans contrainte devient flexible et peut atteindre différentes configurations inactives.




 boucle de liaison au magnésium).
boucle de liaison au magnésium).