Domaine catalytique de la PKA
(Figure : vetopsy.fr d'après Meharena)
Selon ce modèle, l'activation complète des PK nécessite l'assemblage de deux axes ou " colonnes vertébrales " (" spines "), spine C (catalytique) et spine R (régulatrice) pour maintenir une ossature correcte de la kinase afin de sécuriser la position de l'ATP.
Chaque spine comprend plusieurs résidus non polaires situés dans différentes régions qui forment une région hydrophobe continue.
Les deux spines traversent le noyau catalytique et se connectent mutuellement via l'hélice αF, formant ainsi l'intérieur hydrophobe du domaine kinase.
Spine C
1. La spine C contient le cycle de l'adénine de l'ATP pour connecter les résidus hydrophobes des lobes N- et C-terminaux.
Le segment supérieur est formé par deux résidus du lobe N, i.e. Val57 de β2 et Ala70 de β3.
Le segment inférieur est formé par plusieurs résidus du lobe N, i.e. Leu173 de β7 et ses voisins Leu172 et Ile174 qui couvre Met128 de αD, Leu227 et Met231 de αF.
Ces liaisons permettent de caler le squelette de l'ATP, i.e. l'hétérocycle adénine/ribose.
2. La spine C se dissocie lors de la libération de l'ADP et se reconstitue lors d'une nouvelle liaison à l'ATP lors de la conformation ouverte de la PKA.
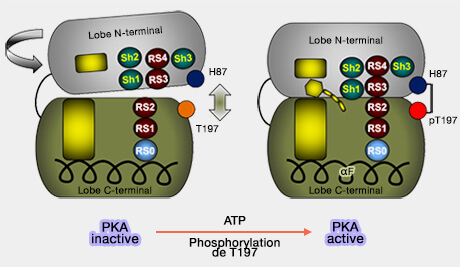
3. La spine R est maintenue dans le lobe N-terminal par trois "coquilles" (shell) formées par un résidu hydrophobe :
SH1 (Val104),
SH2 (Met120),
SH3 (Met118).
Les spines C et R de PKA
(Figure : vetopsy.fr d'après Meharena)
Met120, localisée entre les spines, contrôle l'accès à la poche de liaison liaison, i.e. il est aussi appelé gardien de la porte ou portier (" gatekeeper ").
Selon la PK, ce gardien peut avoir une chaîne latérale volumineuse (comme Phe dans CDK2 et Ck2α) et entraver la pénétration du ligand, ou réduite (comme l'EGFR) pour augmente l'accessibilité du site de liaison
4. Le maintien de l'ossature de la spine R est essentielle à l'activité catalytique de la PKA et nécessite plusieurs interactions indispensables :
l'interaction entre RS1 et RS2, ainsi que celle entre RS0 et l'hélice αF,
l'ancrage de RS3 (hélice αC) par SH1,
la présence de SH3 et SH2 : l'absence des deux abolit l'activité catalytique alors qu'une absence de l'une ou l'autre la diminue.
La perte de l'activité catalytique peut être restaurée par la phosphorylation de la boucle d'activation, créant ainsi la liaison pT197-H87 qui stabilise la conformation de la spine R.
Il semblerait que pT197 interagit aussi avec M189 et T195 de la boucle d'activation, R195 dans la boucle catalytique.
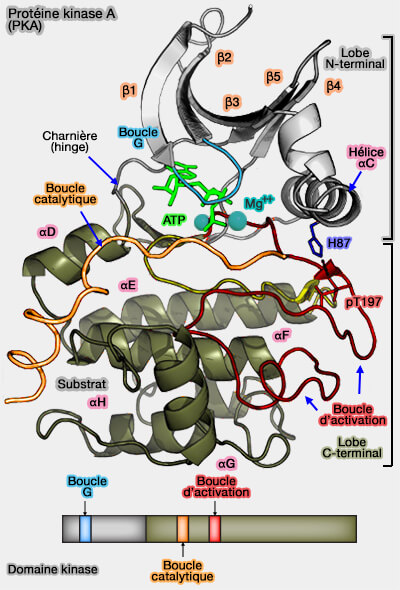
Mécanisme catalytique
1. La phosphorylation de la Thr197 provoque une nouvelle organisation du segment d'activation pour bien placer les ions phosphates en faisant intervenir :
Mécanisme catalytique de la PKA
(Figure : vetopsy.fr) Asp166,
les ions Mg++, qui augmente la positivité du γ-phosphate en neutralisant la charge négative des atomes d'oxygène,
2. L'hélice αC se retourne pour terminer la formation des spines et la bloque simultanément par le contact polaire K72–E91 et le mécanisme catalytique peut s'enclencher.
a. Le résidu sérine/thréonine du substrat est orienté, par des contacts entre l'oxygène du phosphate β et l'ion Mg++, pour que son groupe hydroxyle soit en face du γ-phosphate de l'ATP.
b. Lors de la liaison du substrat, Asp166 de la boucle catalytique agit comme une base et accepte le proton délivré par le groupe hydroxyle de la sérine/thréonine du substrat peptidique.
c. Ce processus déclenche l'attaque nucléophile de l'oxygène chargé négativement sur le phosphate γ via une réaction nucléophile SN2 pour former un complexe transitoire.
d. Puis, le γ-phosphate stabilisé par Mg++ est transféré sur le groupe alcool du substrat de la sérine/thréonine par clivage de la liaison phosphodiester entre les phosphates β et γ pour former une phosphosérine/phosphothréonine et de l'ADP.
e. La moindre affinité de l'ADP pour la poche de liaison du nucléotide l'en fait sortir en étnat déplacé par une autre molécule d'ATP.




 boucle de liaison au magnésium).
boucle de liaison au magnésium).