


Enzymes
Structure en sandwich α-β-α
P-loop versus pli de Rossmann
- Biochimie
- Chimie organique
- Bioénergétique
- Composition de la matière vivante
- Composés organiques
- Protides
- Acides nucléiques
- Glucides
- Lipides
- Enzymes
- Vue d'ensemble
- Classification des enzymes
- Oxydoréductases (EC 1)
- Transférases (EC 2)
- Hydrolases (EC 3)
- Protéases (peptidases ou enzymes protéolytiques)
- Phosphatases
- Estérases
- Lipases
- Phospholipases
- ATPases
- Vue d'ensemble et classification des ATPases
- ATPases à moteur rotatif : F/V/A ATpases
- P-ATPases
- Vue d'ensemble et classification des P-ATPases
- P4-ATPases
- Protéines AAA et AAA+
- Transporteurs ABC
- E-ATPases
- GTPases
- Lyases ou synthases (EC4)
- Isomérases (EC5)
- Ligases ou synthétases (EC 6)
- Coenzymes
- Vitamines
- Hormones
- Composés inorganiques
- Composés organiques
- Transport membranaire
- Moteurs moléculaires
- Voies de signalisation

La structure en sandwich α-β-α est l’un des plus anciens, sinon le plus ancien repliement protéique connu décliné en deux versions, P-loop au sens large et pli de Rossmann.
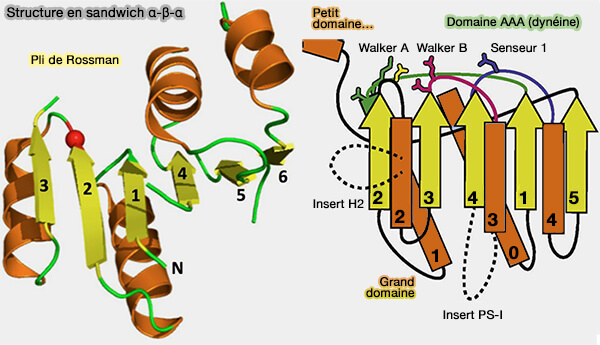
(Figure : vetopsy.fr d'après plusieurs sources)

Vous pouvez lire : On the emergence of P-Loop NTPase and Rossmann enzymes from a Beta-Alpha-Beta ancestral fragment (2020).
Les deux lignées des P-loop des NTPases et des plis de Rossmann sont largement diversifiées et chacune est associée à plus de 120 familles et 75 réactions enzymatiques différentes.
- De plus, ces deux lignées sont omniprésentes dans l’arbre de la vie et l'on considère que ces enzymes sont apparues bien avant le dernier ancêtre commun universel (LUCA) et qu’elles faisaient partie des toutes premières, sinon de la première famille d’enzymes.
- En effet, les boucles P et les boucles de Rossmann sont toutes deux surnommées domaines de liaison aux nucléotides car elles utilisent toutes deux des ribonucléosides phosphorylés tels que l’ATP ou le NAD, ainsi que d’autres cofacteurs pré-LUCA tels que SAM.
Remarque importante : souvent, quand on utilise le terme de P-loop, on fait référence à la boucle de liaison au phosphate (PBL ou Phosphate binding loop) indistinctement pour les NTPases à boucle P, i.e. le motif A de Walker, et celle du pli de Rossmann.
Topologie β
1. Les protéines sandwich αβα sont constituées d’une répétition en tandem d’éléments α-boucle-β, où les boucles supérieures forment le site actif.
- La boucle P minimale ou le pli minimum de Rossmann comprend cinq éléments α-boucle-β reliés par de courtes boucles de connexion ou inférieures qui n’ont généralement aucun rôle fonctionnel.
- Le pli de Rossmann canonique possède 6 brins β.
Remarque : la figure ci-dessous montre un pli de Rossmann à 5 brins pour mieux le comparer à la P-loop.
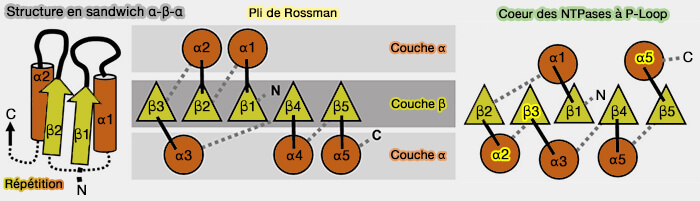
(Figure : vetopsy.fr d'après Longo et coll)
2. Cependant, bien que le pli global soit conservé, la topologie, en particulier l’ordre des brins du feuillet β, diffère entre les plis de Rossmann et les P-loops.
- La topologie du pli de Rossmann, i.e. β3-β2-β1-β4-β5 possède un axe de pseudo-symétrie d'ordre 2 entre β1-β3 et β4-β5 ou β1-β3 et β4-β6 dans les domaines communs à 6 brins (Proteopedia: Rossmann fold: A beta-alpha-beta fold at dinucleotide binding sites 2015)
- La topologie de la boucle P échange au moins deux brins et les boucles P peuvent adopter plusieurs topologies différentes, dont la plus courante est β2-β3-β1-β4-β5.
Liaison aux nucléotides phosphorylés
Similitudes
La deuxième caractéristique commune est que les boucles P et les boucles du pli de Rossman se lient aux ligands ribonucléosidiques phosphorylés en tant que substrats, cosubstrats ou cofacteurs qui sont des phospho-ligands.
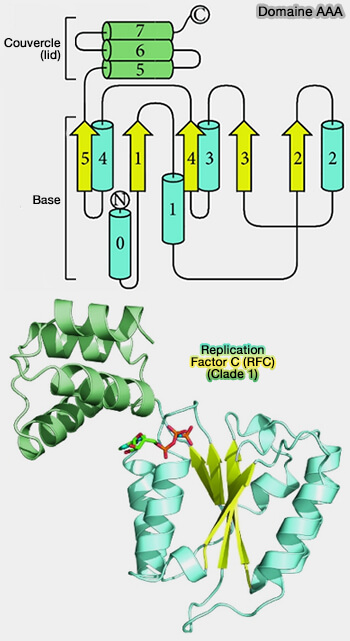
(Figure : vetopsy.fr d'après Miller et Enemark)
1. Bien que le mode global de liaison des ligands diffère, les modes de liaison de leurs fractions phosphate partagent des similitudes.
- Le phosphate est lié par le premier élément β1-loop-α1 qui réside au centre du domaine.
- Les deux boucles de liaison au phosphate médient la liaison via des liaisons hydrogène formées par les amides du squelette à l’extrémité N-terminale de la première hélice α canonique (α1) ainsi que via les résidus de la boucle elle-même.
2. Les deux boucles de liaison au phosphate sont des séquences riches en glycine avec des motifs similaires :
- le motif canonique de Rossmann est GxGxxG,
- le motif canonique de la boucle P, appelé motif A de Walker, est GxxGxGK[T/S].
Différences
1. Cependant, malgré des éléments de liaison au phosphate similaires, le mode de liaison des phospho-ligands des P-loops et des plis de Rossmann est fondamentalement différent.
a. Ils utilisent des nucléotides phosphorylés différents.
- Les boucles P catalysent principalement le transfert du phosphoryle et fonctionnent donc le plus souvent avec l’ATP et le GTP à l’aide d’une dication métallique comme Mg++.
- Les plis de Rossmann utilisent principalement le NADP, les fractions phosphate ne servant que de liaison, tandis que la réaction d'oxydoréduction se produit ailleurs, par exemple, par la base nicotinamide dans le NAD..
b. Ces différences fonctionnelles s’accompagnent d’un certain nombre de différences structurelles dans le mode de liaison du phosphate.
- La boucle P, i.e. motif A de Walker, est une boucle relativement longue, exposée à la surface, qui s’étend au-delà du noyau de la protéine et s’enroule, comme la paume d’une main, autour des fractions phosphate du ligand.
- La PBL du pli de Rossmann est courte et forme une surface d’interaction plane, les groupes phosphate interagissant principalement avec l’extrémité N-terminale d'α1 via une molécule d’eau hautement conservée.
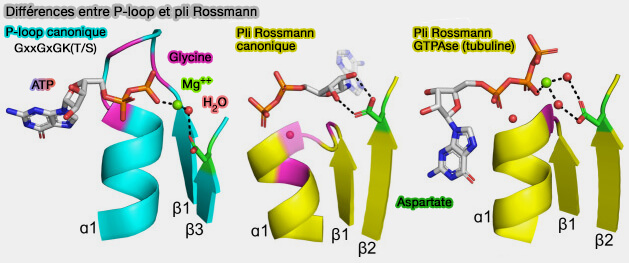
(Figure : vetopsy.fr d'après Longo et coll)
2. L’orientation du phospho-ligand lié est différente.
- La fraction nucléosidique de Rossmann est orientée " à l’intérieur ", i.e. dans la direction du noyau du feuillet β.
- Dans les boucles P, elle pointe " à l’extérieur ", i.e. à l’opposé de l’intérieur de la protéine avec une rotation d’environ 180 ° par rapport au pli de Rossmann.
3. Cette différence d’orientation est liée aux caractéristiques des interactions avec des parties des ligands ribonucléotidiques autres que leurs fractions phosphate.
a. Dans le site de liaison canonique de Rossmann, la fraction phosphate et la fraction ribose sont liées.
- Le phosphate interagit avec le PBL de Rossmann à l’extrémité N-terminale de α1.
- La fraction ribose, forme une interaction unique avec les hydroxyles 2' et 3', est maintenue en place par un résidu Asp/Glu à l’extrémité de β2 (An Ancient Fingerprint Indicates the Common Ancestry of Rossmann-Fold Enzymes Utilizing Different Ribose-Based Cofactors 2016).
b. Dans les boucles P, en revanche, le cœur du domaine αβα n’interagit pas avec le ribose, mais effectue plutôt des interactions plus étendues, orientées catalytiquement, avec les fractions phosphate, via le motif A de Walker et d’autres résidus clés.
- La liaison au phospho-ligand implique également la coordination d’un cation métallique, principalement Mg++, mais aussi Ca++, par deux résidus clés conservés : l’hydroxyle de la sérine canonique ou la thréonine du motif A de Walker (GxxGxGK[S/T]) et un Asp/Glu résidant à l’extrémité d’un brin β adjacent.
- Ce résidu Asp/Glu est au cœur du motif B de Walker, qui est généralement situé à l’extrémité de β3 ou β4 (Evolution of cation binding in the active sites of P-loop nucleoside triphosphatases in relation to the basic catalytic mechanism 2018).
Et le pli Rossmann-like ?
Certains font référence au pli Rossmann-like qui est une variante structurale du pli Rossmann, i.e. il possède un cœur constitué d'un feuillet β entouré d'hélices α avec quelques différences quant :
- au nombre de brins β, leur ordre topologique et leur orientation, parfois antiparallèle,
- aux boucles ou hélices intermédiaires.
On le trouve dans plusieurs familles, dont la superfamille ASKHA (Acetate and Sugar Kinase/Hsc70/Actin fold) dans laquelle les deux domaines actine-fold ou kinase-fold peuvent être considérés comme des plis Rossmann-like ( domaine protéine kinase).
domaine protéine kinase).