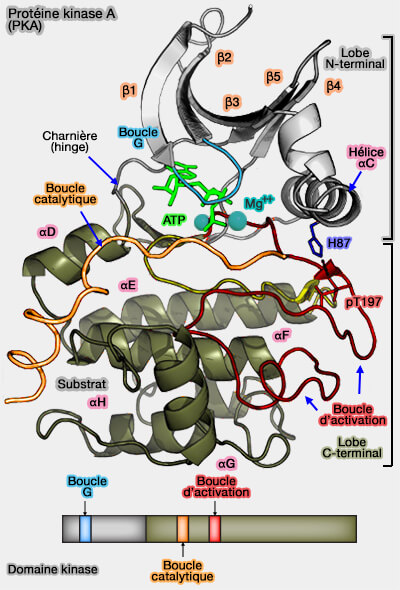
Le lobe C-terminal comprend plusieurs régions essentielles à l'activité catalytique :
la boucle catalytique,
la boucle de liaison au magnésium qui coordonne le complexe Mg2/ATP complexe,
la boucle d'activation qui contient Thr197 dont la phosphorylation est nécessaire à l'activation du domaine catalytique de la PKA,
la boucle P+1 et son environnement pour former le site de liaison peptidique.
Les boucles de liaison, d'activation et P+1 forment le segment d'activation qui possède à la fois une fonction catalytique et régulatrice
Boucle catalytique
La boucle catalytique (catalytic loop), résidus 166-171, entre β6 et β7, contient le motif HRD/YRD.
1. Arg165 déclenche l'assemblage du centre actif en réponse à la phosphorylation de la boucle d'activation et Tyr164 forme le RS1 de la PKA.
2. Asp166 :
accepte le proton délivré par le groupe hydroxyle de la sérine/thréonine du substrat peptidique.
coordonne le groupe hydroxyle de Ser/Thr du substrat au phosphate γ de l'ATP, dont la position est aussi contrôlé par Lys168 par les interactions de Thr197–Arg165–Asp166–Thr201–Lys168.
La position de la boucle catalytique est stabilisée par la liaison entre :
Centre catalytique de PKA
(Figure : vetopsy.fr d'après Kornev et coll)
Segment d'activation
Le segment d'activation est bordé par :
les brins β7/β8,
le motif APE et l'hélice αF.
Le segment d'activation est un polypeptide flexible qui permet les changements conformationnels de l'hélice αC, le motif DFG et la boucle catalytique pour activer le domaine catalytique de la kinase.
Dans la conformation inactive ou fermée de la PKA, elle empêche l'ATP et/ou le substrat de se lier à la PKA.
Dans la conformation active ou ouverte de la PKA, elle oriente et coordonne l'ATP et la reconnaissance du substrat.
Boucle de liaison au magnésium
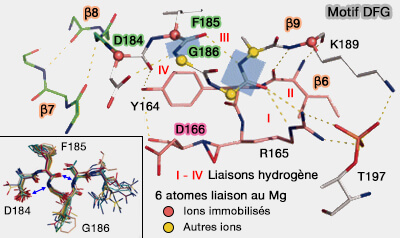
1. La PKA est liée à deux ions magnésium, appelés Mg1 et Mg2, par une petite boucle, résidus 184-187 situés entre β8/β9, qui comprend le motif DFG (Asp184-Phe185-Gly186).
Dans sa conformation active, l'hélice αC se tasse contre le lobe N-terminal et Asp184 du motif DFG chélate les ions Mg++ pour les placer dans le site actif au dessus de l'ATP.
Motif DFG
(Figure : vetopsy.fr d'après Kornev et coll)
Mg1 se lie avec Asp184 (et Asn171), pour positionner les α- et γ-phosphates de l'ATP.
Mg2 se coordonne avec un atome d'oxygène d'Asp184 pour coordonner les β- et γ-phosphates d'ATP.
Les ions Mg++ (ou Mn++ pour d'autres kinases) sont essentiels pour neutraliser la forte charge négative des phosphates et pour coordonner le groupe triphosphate dans le centre actif.
2. Phe185 occupe une poche hydrophobe (DFG-in) adjacent au site ATP et contribue à RS2 ( spine R).
Arg165 de la boucle catalytique, qui déclenche l'assemblage du centre actif,
Lys189 qui permet la formation de formation de deux brins β (β6 et β9) et déclenche la rotation de la liaison peptidique de DFG, i.e. stabilisant davantage Arg165 par des liaisons hydrogène supplémentaires.
Les résidus acides (328-334) de l'extrémité C-terminale augmentent l'attraction des peptides chargés positivement vers le site actif ( extrémité C-terminale)
Glu208 forme une liaison saline avec Arg280 de la boucle αH-αI qui stabilise la position du segment d'activation par rapport aux hélices αG-αI C-terminales.
Ala206 et Pro207 forme un site hydrophobe pour Trp222 de l'hélice αF.
3. En outre, l'intégration croissante entraîne la formation obligatoire d'une liaison entre Lys168 et Thr201, Lys168 coordonnant déjà le groupe hydroxyle de Ser/Thr du substrat au phosphate γ de l'ATP, par les interactions de Thr197–Arg165–Asp166–Thr201–Lys168.
Hélice αF et spines C et R
L'hélice αF, résidus 218-233, est placée au milieu du lobe C-terminal (comme l'hélice αE) sert d'échafaudage pour l'assemblage des spines hydrophobes.



 Boucle de liaison au magnésium).
Boucle de liaison au magnésium).