La région F1 de l'ATP synthase, qui se trouve dans la matrice mitochondriale et qui est responsable de l'hydrolyse de l'ATP, est composée par un ensemble de 5 sous-unités, i.e. α, β, γ, δ et ε.
Structure dimérique de l'ATP synthase bovine
(Figure : vetopsy.fr d'après Spikes et coll)
La région F1 de l'ATP synthase, qui se trouve dans la matrice mitochondriale et qui est responsable de la synthèse/hydrolyse de l'ATP, est composée par un ensemble de 5 sous-unités, i.e. α, β, γ, δ et ε, de stoechiométrie (α)3, (β)3, (γ)1, (δ)1 et (ε)1.
Sites de liaison des nucléotides : sous-unités α et β
Les sous-unités α et β sont alternées et forment un hexamère globulaire d'environ 100 Å de diamètre, i.e. (αβ)3, avec 6 sites de liaison.
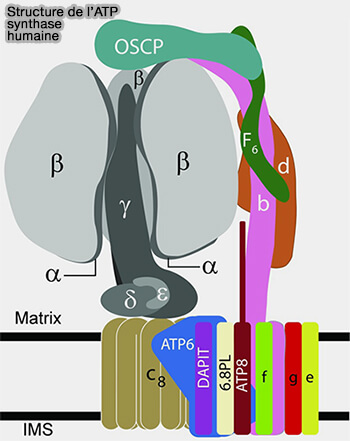
Structure de l'ATP synthase humaine
(Figure : vetopsy.fr d'après Lai et coll)
1. Les sous-unités α et β, presque identiques structurellement, sont constituées par plusieurs domaines.
a. L'extrémité N-terminale est composée par un tonneau β (β-barrel) à 6 brins (a-f).
Les domaines du tonneau chapeautent (αβ)3 par un feuillet à 24 brins.
Aux extrémités N-terminales, 18 à 24 résidus au niveau des sous-unités α et 4 à 8 au niveau des sous-unités β sont désordonnés.
b. Une unité centrale contient le site de liaison au nucléotide, séparé du domaine précédent, mais légèrement interdigité avec le suivant.
Le domaine de liaison aux nucléotides contient un feuillet β à neuf brins associé à neuf hélices α associées.
Les sites de liaison aux nucléotides se trouvent aux interfaces entre les sous-unités α et β.
c. L'extrémité C terminale comprend un faisceau de 7 hélices pour α et 6 hélices pour β.
2. Les trois unités α sont catalytiquement inactives.
a. Par rapport aux sous-unités β, il n'y a pas d'équivalent spatial de Glu188, qui est remplacé par Gln208, la chaîne latérale pointant à l'opposé du phosphate terminal.
La liaison de l'adénine dans la sous-unité α implique plusieurs liaisons hydrogène, tandis que dans la sous-unité β, l'adénine est en contact avec une interface hydrophobe.
Avec la présence de Tyr368 à la position 2 de l'anneau adénine dans la sous-unité α, cela peut expliquer la spécificité des sites nucléotidiques de la sous-unité α pour les nucléotides d'adénine, et la capacité des sites nucléotidiques de la sous-unité β à accueillir l'ATP6.
Structure des sous-unités α et β de l’ATP synthase
(Figure : vetopsy.fr d'après Abrahams et coll)
b. Les nucléotides présents dans les sous-unités α varient selon les auteurs.
3. Les trois unités β catalysent la synthèse de l'ATP, mais sont différentes.
βE, à conformation ouverte, est vide.
βTP, à conformation serrée (tight), contient un MgATP
βDP, à conformation lâche (loose), contient un MgADP et un Pi.
Remarque : l'ion Mg++ est nécessaire à la fixation du nucléotide car il neutralise les charges négatives des groupes phosphate, ce qui stabilise la molécule et lui permet de se lier efficacement au site de fixation du nucléotide.
Sans Mg++, la liaison est instable ou absente car les sites de liaison ne reconnaissent pas le nucléotide seul.
En outre, il est nécessaire à l’attaque nucléophile de l’ADP sur Pi pour former l’ATP dans les sous-unités β.
Structure des sous-unités α, β et γ de l’ATP synthase dans les trois stades
(Figure : vetopsy.fr d'après Abrahams et coll)
Tige centrale
La tige centrale est formée par les sous-unités γ, δ, et ε qui tournent à l'intérieur de la région F1.
Structure de l'ATP synthase humaine
(Figure : vetopsy.fr d'après Lai et coll)
Une hélice N-terminale 1-45 forme un coiled-coil antiparallèle gauche avec l'hélice C-terminale.
Une longue hélice C-terminale 212-272 émergerait de 15 Å au-dessus de (αβ)3.
Sa structure hélicoïdale forme l'axe central autour duquel sont disposés
(αβ)3.
Son côté convexe pousse l'anneau, motif hélice-tour-hélice à l'extrémité C-terminale de la sous-unité y, appelé charnière pour exposer et vider le site de liaison du nucléotide.
La sous-unité γ permet à β de passer par des changements conformationnels, i.e. états fermés, à moitié ouverts et ouverts, qui permettent à l'ATP d'être lié et libéré une fois synthétisé.
Le stockage transitoire d'énergie élastique au cours des paliers rotatifs de 120 degrés de l'enzyme serait dépendant de cette structure qui paraît rigide, mais qui semble possèder des points de flexion au-dessus et en-dessous.
2. La partie globulaire de la sous-unité γ, entre les deux hélices, fait saillie au-dessous de 30 Å, entre en interaction avec les sous-unités δ et ε, et fixe F1 fermement à la figure du domaine FO.
Sous-unité δ/ε
1. La sous-unité δ au niveau mitochondrial, qui correspond à la sous-unité ε au niveau des bactéries et des chloroplastes, renforce l'attachement de la sous-unité γ aux sous-unités c de FO.
une hélice α domaine qui contient des segments hélice1/boucle2/hélice2/crochet qui sont censés changer les conformations entre une forme étendue et une forme compactes.
2. Dans les mitochondries, on trouve en plus de δ, une sous-unité ε, terme mal choisi, qui renforce cette attache.
Les sous-unités γ/δ fixent fortement la structure cylindrique hydrophobe constituée d'une couronne de sous-unités c du domaine membranaire FO de l'enzyme et forment ensemble le rotor de l'enzyme.
Tige périphérique (PS)
La tige périphérique (PS), i.e. peripheral stalk, est stationnaire et est constituée :
au niveau de F1 par la sous-unité OSCP (Oligomycin Sensitivity Conferral Protein) des mitochondries qui correspond à la sous-unité δ des bactéries et des chloroplastes,
au niveau de FO par les sous-unités F6, b, d et la région extramembranaire d'ATP8/A6L i.e. b et b' des bactéries et des chloroplastes.
Attention de ne pas confondre l'OSCP à sous-unité δ !
La tige périphérique relie la surface externe du domaine catalytique à la sous-unité a/ATP6 dans le domaine membranaire.
Structure de la tige périphérique de l'ATP synthase
(Figure : vetopsy.fr d'après Dickson et coll)
a. Son domaine N-terminal globulaire de six hélices (H1-H6) coiffe le sommet du module catalytique F1.
Il fournit le site de liaison, par les hélices H1 et H5, pour la région N-terminale d'une de trois sous-unités, qui s'étendent au-dessus du domaine (αβ)3, en particulier αE, sous-unité α vide.
Dans l'article sur la structure de l'ATP synthase bovine, il est indiqué qu'elle s'attache sur les 3 sous-unité α ( figure S19).
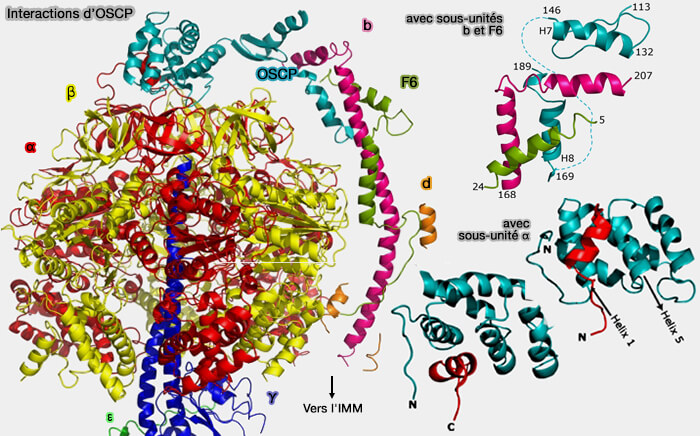
Interactions d'OSCP
(Figure : vetopsy.fr d'après Rees et coll)
b. Son domaine C-terminal est constitué d’une structure plus étendue composée de deux longues hélices (H7 et H8) séparées par des feuillets β parallèles et antiparallèles consécutives, i.e. β1-β2 et β3-β4 ( figure S20-22).
Il interagit avec les sous-unités b et F6 de la tige périphérique.
Cette flexibilité serait principalement due au résidu de glycine G114 situé dans la région de la charnière.
La flexibilité de la tige périphérique lui permet de s'adapter aux mouvements verticaux et transversaux du domaine α3β3 pendant la catalyse causée par le rotor asymétrique ( rôles de la tige périphérique).
Structure de l'ATP synthase humaine
(Figure : vetopsy.fr d'après He et coll)
2. OSCP est attachée à une tige rigide de 150 Å de longueur formée par des portions parallèles de régions α-hélicoïdales ( figure S4 et 23) :
de la sous-unité b, i.e. résidus 91 à 186 et 190 à 214, sous-unité formant une hélice α continue et incurvée d’environ 160 Å de long ( partie membranaire de la sous-unité b).
de la sous-unité d, i.e. résidus 24 à 43, 53 à 59, 63 à 75, 84 à 122 et 132 à 138, sous-unité constituée de 5 hélices α-hélicoïdales.
de la sous-unité F6, i.e. résidus 9 à 24 et 33 à 50, sous-unité constituée de 2 hélices α-hélicoïdales, renforcée par la région C-terminale de ATP8/A6L, i.e. résidus 30 à 66.
Remarque : chez les bactéries, le domaine N terminal de la sous-unité δ est similaire à celle d'OSCP.
Les deux sous-unités b bactériennes, i.e. 2 copies de la même sous-unité b ou 2 protéines apparentées, b et b', qui correspondent aux sous-unités I et II dans les chloroplastes, forment un domaine coiled-coil.
3. La tige rigide est connectée au domaine membranaire du stator, i.e. domaine a/ATP6, par une charnière de la sous-unité b, i.e. résidus de 73 à 90.




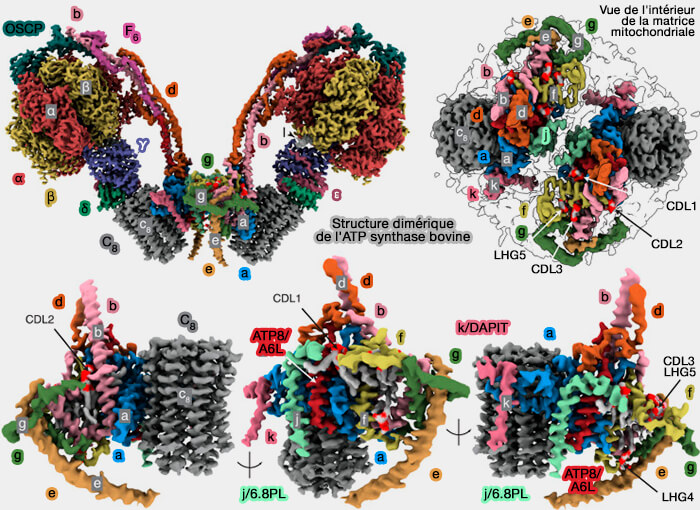

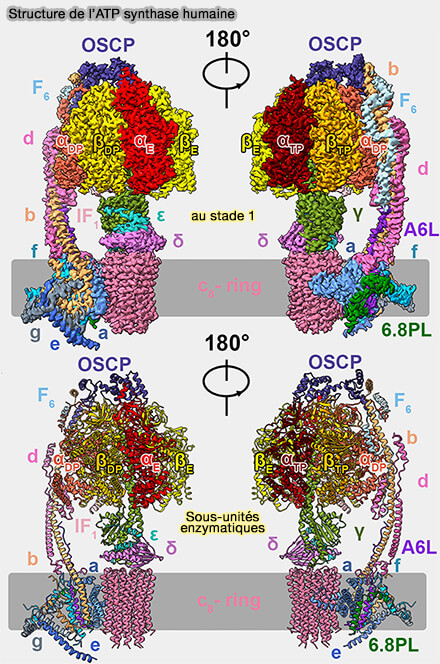
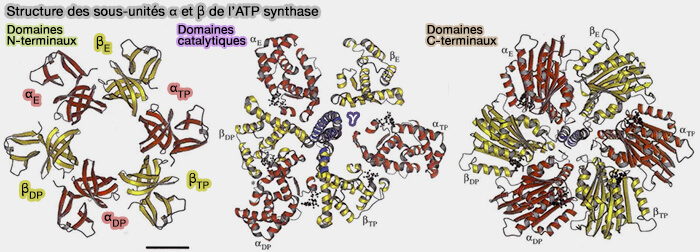
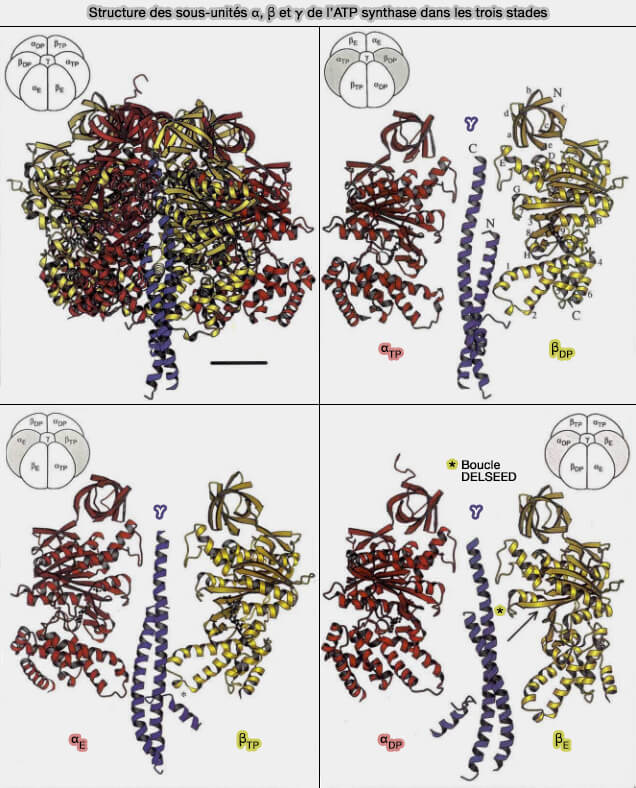
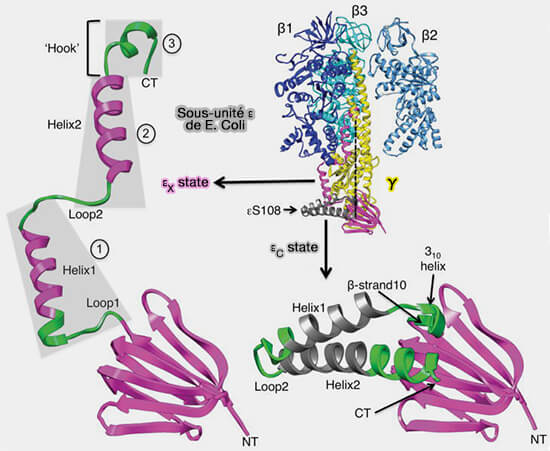

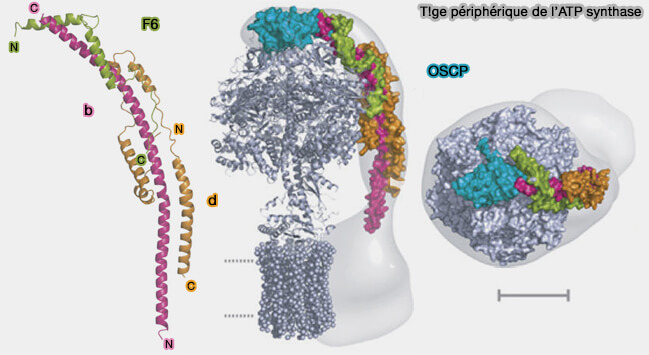
 figure S19).
figure S19).