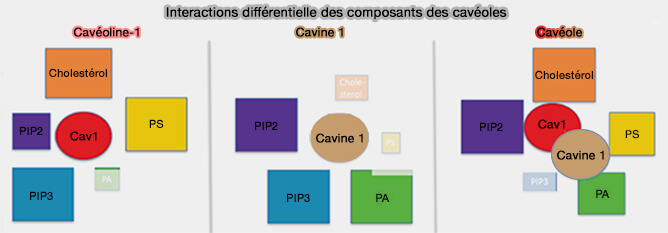
Les cavéolines sont nécessaires aux cavines pour former des cavéoles, en s'oligomérisant et en s'associant aux phospholipides membranaires, i.e. elles forment un complexe de revêtement externe qui remodèle la membrane en cavéoles.
Vue d'ensemble de l'oligomérisation
L'oligomérisation est produite par de vastes réseaux d'interactions tout le long de chaque protomère de Cav1, y compris les régions du bord, du rayon et du moyeu.
Complexe Cav1 8S et oligomérisation
(Figure : vetopsy.fr d'après Porta et coll)
1. En partant du bord, l'extrémité N d'un protomère (i) engage quatre protomères voisins, i.e. i - 2, i - 1, i + 1 et i + 2.
Les OD des protomères voisins sont disposés les uns à côté des autres, mais ne se chevauchent que légèrement.
Interactions des protomères de Cav1
dans le complexe Cav1 8S
(Figure : vetopsy.fr d'après Porta et coll)
Dans ODi, SMi est pris en sandwich entre α1i + 1, α2i + 1, α2i + 2, α3i + 2, π1i + 2 et PMi + 1.
Arg54i + 1 de PMi + 1 s'intercale dans une poche formée par His79i et Trp85i, épinglant ODi en place jouant un rôle majeur dans la formation du complexe 8S.
2. Les hélices de l'IMD hautement hydrophobes (α2, π1 et une partie de α3) contribuent à l'oligomérisation, en particulier la chaîne latérale de Trp128i + 2.
3. Les hélices du SR (partie C-terminale de α3, α4 et α5) établissent des contacts par paires entre les protomères pour finalement former le tonneau β.
Ils impliquent des résidus avec des chaînes latérales plus grandes, augmentant la séparation des hélices par rapport à celles du croisement α1i-α2i + 1, qui convergent finalement pour former le tonneau β C-terminal, la région majeure finale des interactions protomère-protomère.
Complexe 8S et modèle en orange
(Figure : vetopsy.fr d'après Han et coll)
Modèle des interactions des cavéolines
Interactions avec la membrane
Modèle dominant
1. Le modèle dominant suppose que l'IMD de Cav1 forme une structure en épingle à cheveux qui s'insère dans la membrane, créant un coin qui plie la membrane (Key Phases in the Formation of Caveolae 2021).Cavéoline-1 et interactions membranaires
(Figure : vetopsy.fr d'après Busija et coll)
2. La cavéoline pourrait être insérée dans la membrane dans deux états interconvertibles distincts ( cavéoles et cholestérol).
Le premier état, i.e. insertion partielle, n'est capable de générer qu'une faible courbure et se caractérise par une insertion moins stable dans la membrane, i.e. comme la cavéoline dans le complexe de Golgi ou dans la membrane plasmique des cellules appauvries en cholestérol.
Un deuxième état, i.e. insertion complète, avec une conformation plus entièrement insérée dans la membrane, serait associée à une courbure plus élevée de la membrane et à la scission des vésicules, i.e. comme dans les membranes d'E. Coli ou les membranes mammaliennes enrichies en cholestérol par leur laison avec le SD.
Cavéoles et phospholipides
(Figure : vetopsy.fr d'après Zhou et coll)
La structure du complexe 8S Cav1 montrerait que l'IMD contribue à la formation d'une surface plane faisant face à la membrane tout en stabilisant simultanément les contacts entre les protomères.
Dans ce modèle, le côté du complexe associé à la membrane s'intègre profondément dans le feuillet cytoplasmique, interagissant avec les carbones terminaux des lipides du feuillet opposé plutôt que de s'asseoir à l'interface entre les groupes de tête et les chaînes acyle, comme serait généralement attendu pour les hélices amphipathiques.
Le complexe 8S, en déplaçant les lipides du feuillet cytoplasmique, pourrait créer un nanodomaine membranaire ordonné composé de protéines sur un feuillet membranaire et de lipides sur l'autre.
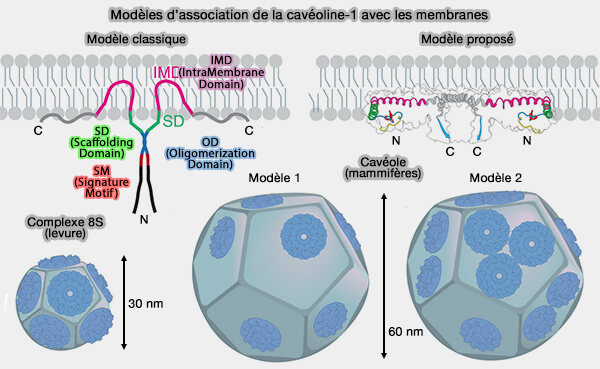
Modèles d’association de la cavéoline-1 avec les membranes
(Figure : vetopsy.fr d'après Porta et coll)
Association des complexes Cav avec la membrane
Plusieurs modèles peuvent être décrits pour l'association des complexes 8S avec les membranes.
1. Un seul complexe 8S s'adapte parfaitement sur chaque face d'un dodécaèdre de la taille caractéristique des h-cavéoles de 30 nm de diamètre moyen qui devrait contenir 132 copies de Cav1, i.e. 144 ± 39 copies par cavéole (Molecular Characterization of Caveolin-induced Membrane Curvature 2015).
2. Cependant, comme une cavéole de mammifère a un diamètre moyen de 61 nm en supposant une symétrie dodécaédrique, il se peut que :
soit une partie de chaque face est occupée par des protéines supplémentaires.
Les complexes de cavéoline ont pour rôle principal de stabiliser les surfaces membranaires plates des structures polyédriques, plutôt qu'en imposant une courbure membranaire continue, i.e. mécanisme encore à définir pour les protéines membranaires intégrales, de modifier les membranes cellulaires pour former des domaines fonctionnels.








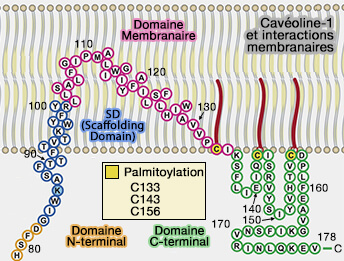
 cavéoles et cholestérol).
cavéoles et cholestérol).